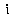

Физиология пищеварения
С. С. Полтырев‚ И.Т. Курцын (1980 год)
В пособии содержатся данные, характеризующие состояние органов пищеварения при разных температурах окружающей среды, на высотах, при беременности и лактации, болевых ощущениях, умственной и мышечной деятельности. Рассмотрены вопросы гигиены питания детей и подростков, заболевания пищеварительных органов, возникающие вследствие нарушения режима питания. Дано представление о функциональной связи пищеварительной системы с другими системами организма и механизме ее осуществления. Книга может быть полезна специалистам-физиологам, врачам, эндокринологам.
Предисловие
Пособие написано с учетом раздела «Физиология пищеварения» современных программ курсов «Физиология человека и животных», «Возрастная физиология», «Школьная гигиена», в которых основное место занимает физиология пищеварения как начальный этап обмена веществ, оставляющий основу жизни животного организма. Этот раздел физиологии, созданный Иваном Петровичем Павловым, получил дальнейшее развитие в трудах его учеников и последователей как в СССР, так и за рубежом.
Книга знакомит студентов с историей развития физиологии пищеварения в нашей стране и за рубежом; современным состоянием учения о работе органов пищеварения в норме и при патологии у взрослых и детей; вопросами гигиены питания; современными методами исследования пищеварения у человека и животных в эксперименте. Большое место занимают вопросы регуляции деятельности пищеварительной системы.
Все предложения, дополнения, критические замечания, направленные на улучшение настоящего пособия, будут приняты с благодарностью.
Глава 1
История развития физиологии пищеварения
Современный уровень представлений о физиологии пищеварения тесно связан с замечательными исследованиями великого русского ученого Ивана Петровича Павлова (1849—1936) и его школы, сыгравших выдающуюся роль в истории физиологии вообще и физиологии пищеварения в частности.
Исследования И. П. Павлова стали отправным пунктом для создания объективного, строго научного метода изучения деятельности головного мозга, при помощи которого было разработано гениальное учение о высшей нервной деятельности. Они легли в основу современных знаний по физиологии и патологии пищеварения человека и животных, став поистине неисчерпаемой сокровищницей новых творческих дерзаний ученых.
Мыслители и врачи разных времен интересовались ходом пищеварительного процесса. Над ним задумывались философы Аристотель и Декарт, но размышления их носили отвлеченный характер и были чисто умозрительными. Нужно было опытным путем изучить и понять, как пища, поступившая в организм, подвергается перевариванию, превращаясь в конечном счете в клетки и ткани человека и животного. Нужно было узнать, как работают органы пищеварения и какие жидкости они выделяют в просвет желудочно-кишечного тракта, под влиянием которых происходит переваривание сложнейших пищевых веществ и усвоение их организмом. Эти и многие другие вопросы, связанные с процессом пищеварения, должны были привести к выводу о том, что для их выяснения единственно возможным является экспериментальный путь изучения.
В ХVII в. Репе де Грааф изучал слюнную и панкреатическую секрецию у собаки, вводя в протоки околоушной и поджелудочной желез трубочки, однако полученные при этом данные мало обогатили физиологию.
В XVIII в. Спалланцани и Реомюр пытались получить желудочный сок, вводя в желудок привязанную на ниточке резиновую губку. Примитивность методики и неточность проводимого анализа не дали ясного и полного представления о секреторной функции желудка.
В начале ХIХ в. Пру обнаружил в желудочном содержимом наличие соляной кислоты, но не объяснил ее значения для желудочного пищеварения и роли в механизме регуляции деятельности органов пищеварения.
С 1825 по 1833 г. Бомон проводит наблюдения над желудочной секрецией у человека, имевшего осле ранения свищ желудка. Полученные при помощи этих оригинальных но тому времени исследований данные были отрывочны и недостаточно достоверны. При проведении наблюдений Бомон не учитывал исходного функционального состояния железистого аппарата, не принимал во внимание роли влияний коры больших полушарий головного мозга в процессе возбуждения секреторных клеток и не дал точной характеристики количественного и качественного состава желудочного сока. При действии тех или иных пищевых раздражителей. То же самое можно сказать и относительно исследований, проведенных позднее Бушем па человеке со свищом тонкой кишки и Рише — па человеке со свищом желудка.
Отсутствие хорошо разработанных экспериментальных методов исследования приводило в ряде случаев к использованию примитивных приемов наблюдения. Например, Тидеман и Гмелин, а впоследствии Браун с целью выяснения роли механического раздражения в процессе возбуждения желудочной секреции вводили в желудок животных мелкие камни и спустя несколько часов, убив животных, смотрели, имеется ли в желудке сок. Полученные данные были разноречивые и мало убедительны. Подобный прием применяли и другие авторы. Так, Леуре и Лассажне давали собакам проглатывать кусочки губки и через 24 ч, убив животных, исследовали кишечное содержимое, выжатое из губок. Такие методы исследования вели к противоречивым результатам, а порой и к неверным взглядам на функцию пищеварительных желез.
На основании проведенных наблюдений Шульц пищеварительную функцию приписывал только слепой кишке, а Блондло, подробно описывая различные пищеварительные соки, отрицал какое-либо переваривающее действие кишечного и панкреатического соков на пищу. К отрицанию способности кишечного сока переваривать пищу (белок и крахмал) пришел на основании своих опытов и Функ.
В 1842 г. талантливый русский хирург В. А. Басов, наложив фистульную трубку на желудок собаки, впервые предложил новый метод исследования желудочной секреции. Собак с хронической фистулой желудка В. А. Басов и известный русский физиолог А. М. Фило-мафитский демонстрировали студентам во время лекций. Этот оперативный прием, безусловно, сыграл прогрессивную роль в развитии экспериментальной физиологии, но данные, полученные при опытах на животных, оперированных таким образом, отличались большой неточностью, так как деятельность желудочных желез определялась на основании анализа не чистого секрета, отделяемого клетками, а содержимого желудка, состоящего из смеси желудочного сока с пищевой массой.
В 1849 г. Барделебен впервые произвел на собаке операцию перерезки пищевода (эзофаготомию), но ни сам автор, ни последующие физиологи не использовали этот ценный оперативный прием для научно-исследовательских целей.
Ряд исследователей (Флюранс, Гаубер, Колен, Эллен-бергер) применяли фистульный метод на разных сельскохозяйственных животных (коза, овца, корова), но полученные экспериментальные данные были отрывочными и неполными.
В 1852 г. тартуские физиологи Ф. Биддер и С. Шмидт описали психическое отделение желудочного сока у собак, не дав этому явлению объективного, строго научного толкования.
Таким образом, первые экспериментальные исследования по физиологии пищеварения имеют ишь исторический интерес. На основании полученных данных нельзя было построить стройного представления о работе пищеварительного аппарата, физиологической роли органов пищеварения и механизме регуляции их деятельности.
Лишь в Х1Х в. И. П. Павлов с сотрудниками приступил к систематическому изучению физиологии пищеварения. В Западной Европе в это время были широко распространены идеалистические философские взгляды Гегеля и Канта, порождавшие среди естествоиспытателей ненаучные, мистические представления. О состоянии теоретических наук того времени Ф. Энгельс писал:
«Конечным результатом были господствующие теперь разброд и путаница в области теоретического мышления. Нельзя теперь взять в руки почти ни одной теоретической книги по естествознанию, не получив из чтения ее такого впечатления, что сами естествоиспытатели чувствуют, как сильно над ими господствует этот разброд к эта путаница, и что имеющая ныне хождение, с позволения сказать, философия не дает абсолютно никакого выхода». Маркс К, Энгельс Ф. Соч. 2-е изд., т. 20, с. 368. Естественно, то такое состояние теоретических наук не могло не отразиться на методологических принципах и методических приемах физиологов, биологов, патологов и клиницистов.
В этот период времени, невзирая на то, что строение пищеварительной системы уже было остаточно полно описано анатомами, представления о работе отдельных пищеварительных желез и деятельности всего пищеварительного аппарата в целом были весьма скудными и отрывочными.
Аналитический прием исследования был почти единственным при изучении сложнейших физио- логических и патологических явлений. Организм представлялся как сумма отдельных клеток, функционирующих автономно, самостоятельно, вне связи со всем организмом. В физиологии и патологии господствовал вивисекционный метод исследования, т. е. изучение физиологических функций на вырезанных из организма органах и тканях (метод изолированных органов) или на организме, подвергнутом действию наркоза и травмирующих ткани хирургических манипуляций
(метод острых опытов). Естественно, что такой методический прием не мог дать полного и счерпывающего представления ни 0 функции изучаемого органа, ни о связи его с другими органами желудочно-кишечного тракта, ни тем более о физиологических механизмах, регулирующих деятельность органов и всей системы пищеварения в целостном, нормальном организме.
Физиология, по словам И. П. Павлова, «не имела дела со всей полнотой жизненной деятельности организма». Основанная на аналитических данных, она была, по его выражению, «отрывочной», «лоскутной».
Во второй половине ХТХ в. физиологи много работали над изучением деятельности слюнных желез у животных, но, пользуясь в наблюдениях методом острых опытов, сумели выяснить лишь некоторые стороны секреторного процесса без учета роли и значения высших отделов центральной нервной системы — коры головного мозга, а также функции слюнных желез при нормальных физиологических условиях в целостном организме. Р. Клеменциевич и Гейденгайн разработали на собаке операцию наложения изолированного маленького желудочка, перерезая при этом идущие к ему нервные веточки, выключали влияние нервных импульсов на деятельность секреторных клеток, результате чего работа последних во время пищеварения не отражала полной картины пищеварительного процесса в большом желудке с неповрежденной ипнервацией. Тири, а затем Велла разработали операцию изоляции у животного отрезка кишечника с сохраненной ипнервацией. На оперированных таким образом животных удалось выяснить некоторые пищеварительные свойства кишечного сока. И условия, при которых происходит его выделение. Цепные данные о кишечном соке, его ферментативных свойствах были получены А. Доброславпным, В. В. Пашу-тиным и др. Однако, несмотря на большое число экспериментальных работ, во взглядах ученых относительно условий отделение кишечного сока и его физико-химических свойств существовали противоречия и путаница.
Ряд исследователей уделили внимание методам получения желчи и анализу качественного состава ее у животных, но при этом ими строго не соблюдались условия физиологического эксперимента, что, естественно, не дало каких-либо достоверных данных о механизмах регуляции желчеобразовательной желчевыделительной функций печени.
В 1895 г. Фремоп разработал па собаке операцию изоляции всего желудка, подобно тому как это производится с изоляцией кишки по Тири, однако этот способ имел некоторое значение лишь при анализе отдельных звеньев секреторного процесса.
Людвиг, Бернар, Бернштейн, Ландуа и Гейденгайн много работали над изучением функции поджелудочной железы у животных, применяя вивисекцию. Полученные экспериментальные данные были настолько противоречивы и неясны, что сами авторы с горечью и разочарованием вынуждены были заявить о том, что в результате изучения функции поджелудочной железы образовались горы трупов животных, а знания о деятельности этого органа почти не изменились.
Важные данные о физиологии поджелудочной железы были получены студентом Петербургского университета Н. Лебедевым, который впервые доказал, что симпатический нерв является секреторным нервом поджелудочной железы.
Попытки некоторых физиологов перейти к изучению деятельности поджелудочной железы на животном в хроническом опыте путем вживления стеклянной или металлической трубки в проток железы не увенчались успехом, так как спустя несколько дней после операции трубки выпадали.
Следовательно, стремления некоторых естествоиспытателей перейти от вивисекционного метода к методу хронических фистул не дали ощутимых результатов; полученные данные носили подчас случайный, бессистемный характер и в большинстве случаев не отражали истинной картины нормальной деятельности органов пищеварения.
Таким образом, физиологи, экспериментируя в подавляющем большинстве случаев в условиях строго опыта, занимались исследованием функции какого-либо одного органа. В своих поисках они, сходя из принципов аналитической физиологии, изучали деятельность пищеварительного аппарата не как целостной физиологической системы, тесно связанной со всем организмом, а как системы, состоящей из отдельных элементов, взаимно не связанных и работающих изолированно от деятельности всей системы и организма в целом. Их подход к изучению физиологических явлении был метафизичен и абстрактен. Такое положение было характерно не только для биологии и физиологии, но и для медицины начала ХIХ в.
Отсутствие специальных методов исследования функций пищеварительных органов у человека и скудность знаний по физиологии пищеварения не позволяли практическим врачам правильно оценивать работу пищеварительного аппарата при нормальных и особенно болезненных состояниях организма. Некоторым прогрессом в этом направлении явились работы Кусмауля, предложившего методику толстого желудочного зонда, и Лейбе, применившею эту методику для диагностики заболеваний желудка. Позднее Эвальд и Боас с целью анализа желудочной секреции у человека предложили использовать в клинической практике пробный завтрак. Однако методика толстого желудочного зонда страдала рядом существенных дефектов, среди которых особенно важными вились, во-первых, одномоментность взятия пробы желудочного содержимого, и, во-вторых, невозможность анализировать работу желудочных желез по абсолютным показателям секреции, так как полученный сок был смешан с введенным пробным завтраком. Цепные данные о моторике желудка были получены Уффельманом, который при помощи манометрографической методики записывал состояние тонуса и движения желудка у ребенка, имевшего желудочную фистулу.

И. П. Павлов
Таким образом, физиология допавловского периода шла ощупью, накапливая отдельные данные, подчас извращенные и далекие от истины. Сложнейшие физиологические и биохимические процессы трактовались с метафизических позиций. Экспериментаторы не располагали хорошим и надежным методом исследования, у них отсутствовали система и последовательность наблюдения.
Приступая к изучению функций органов пищеварительной системы, И. П. Павлов и его сотрудники вначале воспользовались вивисекционным методом, широко распространенным в аналитической физиологии для решения вопросов физиологии и патологии. Однако вскоре же они убедились в том, «...что обыкновенное простое резание животного в остром опыте, как это выясняется теперь с каждым днем все более и более, заключает в себе большой источник ошибок, так как акт грубо: о на рушения организма сопровождается массою задерживающих влияний на функцию разных органов». Павлов И. П. Поли. собр. соч., т. 11, кн. 2. М., 1951, с. 35.
В поисках НОВЫХ путей исследования И. П. Павлов разрабатывает новый метод — метод хронических фистул, дающий возможность изучать деятельность органов пищеварительного аппарата а целом, неповрежденном животном, при естественных условиях его существования. Это было завершением той замечательной идеи, которой руководствовался В.А, Басов при разработке операции наложения хронической фистулы желудка собаки. Метод хронических фистул явился в дальнейшем основой прогресса экспериментальной физиологии.
Широко применяя в физиологии новый метод изучения, И. П. Павлов и его сотрудники в эпоху развития аналитической физиологии создают новое, оригинальное направление в естествознании — синтетическую физиологию основная идея которой — изучение функции органа в целостном организме, во взаимосвязи его с другими органами и системами. Задача анализа — возможно подробнее ознакомиться с какой-либо изолированной частью и определить отношение ее ко всем возможным явлениям природы. Цель же синтеза состоит в том, чтобы оценить значение каждого органа с истинной и жизненной стороны, указать его место в жизнедеятельности всего организма.
Следовательно, синтетическая физиология предусматривает изучение функции каждого отдельного органа пищеварения в связи с функциями органов и всего организма в целом при нормальных физиологических условиях. Анализ физиологических явлений и их синтез не исключают друг друга, а взаимно связаны и представляют диалектическое единство. Таким образом, единство анализа и синтеза является основным принципом синтетической физиологии. Такой подход к изучению природы принципиально отличался от методов аналитической физиологии, расщепляющей организм а отдельные части и исследующей физиологические процессы в искусственно созданных условиях.
Основные принципы павловского метода изучения работы пищеварительных желез были точно и определенно сформулированы самим И. П. Павловым. «Нужно уметь,— говорил он,— достать реактив во всякое время (курсив наш — С. П., И. К.), иначе бы от нас могли ускользнуть важные моменты, в совершенно чистом виде, иначе мы не будем в состоянии знать изменение состава, нужно очно определять его количество, и, наконец, необходимо, чтобы пищеварительный канал правильно функционировал и животное было вполне здорово». Павлов И П. Поли. собр. соч., т. 11, кн 2. М., 1951, с. 22.
Несмотря на то что большинство органов пищеварения заложено глубоко в брюшной полости и доступ к ним затруднен, И. П. Павлов и его ученики с успехом претворяют в физиологическую практику эти принципиальные положения. При физиологической лаборатории создается специальная операционная, подобная операционной хирургических клиник, где с соблюдением всех условий септики и антисептики производят сложнейшие операции па животных с тем, чтобы после их выздоровления свободно получать чистый секрет пищеварительных желез.
Для экспериментального изучения деятельности органов пищеварения в павловской лаборатории были разработаны и предложены многие методические приемы, в числе которых можно назвать такие, ак наложение постоянных фистул протоков слюнных желез, наложение фистулы желудка в сочетании с эзофаготомией (фистулой пищевода), образование изолированного маленького желудочка сохраненной иннервацией, наложение фистул общего желчного протока и протока поджелудочной
Железы, изоляция различных отделов кишечника. Суть большинства этих операция состоит в том, что из стенки пищеварительного тракта иссекаются отдельные участки с местом впадения протоков той или иной пищеварительной железы и эти участки затем вживляются в рану. Это дает возможность после выздоровления животного получить чистый секрет из открывающихся наружу протоков пищеварительных желез. Особенно следует отметить операции гастроэзофаготомии и образования маленького желудочка. Благодаря сочетанию эзофаготомии с наложением фистулы желудка по Басову, И. П. Павловым и Е. О. Шумовой-Симаповской был поставлен знаменитый опыт «мнимого кормления», который до сих пор является самым демонстративным и оригинальным опытом экспериментальной физиологии. На основании проведенных исследований было показано, что акт еды является сильнейшим возбудителем желудочных желёз и блуждающие нервы являются секреторными нервами желудка.

Маленький желудочек по Павлову широко вошел в обиход всех физиологических лабораторий мира. При помощи изолированного желудочка экспериментатор мог наблюдать, как в зеркале, всю поразительную картину секреторного процесса в большом желудке во время пищеварения.
Разработка новых методических приемов исследования потребовала от И. П. Павлова и его сотрудников много времени и энергии. Так, например, в течение четырех лет Павлов добивался получить животное с хронической фистулой протока поджелудочной железы, и только в 1879 г. после 50 неудачных операций это, наконец, ему удалось.
О преимуществе метода хронических фистул перед вивисекционным методом можно судить по результатам исследования работы некоторых пищеварительных желез в условиях хронического и острого опытов на собаках (табл. 1).
Свыше 25 лет И. П. Павлов с сотрудниками занимался изучением опросов физиологии и патологии пищеварения. Результатом этой напряженной работы явились многочисленные диссертационные работы сотрудников павловской лаборатории и два фундаментальных руда самого И. П. Павлова: «Лекции о работе главных пищеварительных желез» (1897), «Физиологическая хирургия пищеварительного тракта» (1902), сьнравших важную роль в развитии всей мировой физиологии и клиники заболеваний органов пищеварения. Этими трудами была заложена основа современного понимания физиологии пищеварения. На основании огромного количества фактов, полученных в павловской лаборатории, было создано новое представление о ходе и последовательности пищеварения в различных отделах желудочно кишечного тракта; зависимости секреторной реакции пищеварительных желез от сорта пищи, количестве и качестве отделяемого секрета на тот или иной сорт пищи; развитии ферментативных процессов во время пищеварения; приспособлении железистых клеток к длительному качественно различному питанию; характере секреции слюны, желудочного сока, желчи, поджелудочного сока и кишечных соков при действии различных раздражителей; взаимосвязи между отдельными органами пищеварительной системы; двигательной функции желудочно-кишечного тракта, о связи последней с секреторными процессами.
Полученный экспериментальный материал дал не только точную характеристику работы пищеварительного аппарата, но и позволил выяснить основные физиологические механизмы, регулирующие деятельность как отдельных пищеварительных желез, так и всего пищеварительного аппарата в целостном организме, причем была установлена исключительная роль центральных иннервационных механизмов в этом процессе.
Так, со всей отчетливостью и точностью было показано, что при отсутствии соответствующих раздражителей пищеварительные железы (слюнные, желудочные) находятся в состоянии относительного покоя и не выделяют секрета; деятельность желез начинается с момента начала приема пищи или, если последняя действует на животное своим видом и запахом, на расстоянии.
В результате многочисленных наблюдений И. П. Павлов выдвигает и экспериментально обосновывает повое оригинальное учение о пищевом центре, согласно которому регуляция двигательных и секреторных процессов, протекающих во время пищеварения, осуществляется при помощи коры больших полушарий и подкорковых ганглиев.
Основной идеей всех экспериментальных исследований И. П. Павлова и его школы является идея «нервизма», т. е. идея о главенствующей роли нервной системы в регуляции деятельности органов и систем организма. Она пронизывает все исследования великого ученого в области физиологии пищеварения, начиная с одной из его первых экспериментальных работ, посвященной нервной регуляции деятельности поджелудочной железы, и кончая классическим трудом «Лекции о работе главных пищеварительных желез», принесшим И. П. Павлову мировую известность. Идея «нервизма» легла в основу павловских принципов целостности организма и взаимодействия организма с внешней средой. Стало вполне очевидным, что нервная система связывает, во-первых, многочисленные части организма между собой и, во-вторых, организм как сложнейшую систему — с внешней средой.
Значение павловских трудов по физиологии пищеварения исключительно велико. За выдающиеся исследования в этой области И. П. Павлову в 1904 г. была присуждена высшая мировая награда — Нобелевская премия.
Труды И. П. Павлова и его школы открыли новые пути исследования в физиологии и клинике.
Советские ученые, творчески разрабатывая основные принципы павловского учения, добились крупнейших достижений, выдвинувших советскую науку о пищеварении на первое место в мире.
Методологический подход павловской школы к изучению функций органов пищеварения и новое представление о работе пищеварительного аппарата направили совершенно по иному пути развитие клинической мысли и явились решающим фактором в создании новых клинических методов исследования. Воспитанные на замечательных идеях павловского учения, клиницисты стали шире подходить к оценке симптомов заболевании желудочно-кишечного тракта, изменив и уточнив диагностику и лечение и заново перестроив диететику, т. с. учение о питании здорового и больного человека.
И. П. Павлов считал, что конечная и основная задача физиологии — изучить физиологические механизмы, регулирующие деятельность органов и систем всего организма, понять всю сложность динамики жизненных процессов и на основе этого построить правильный и рациональный режим питания, труда и отдыха.
В настоящее время павловская физиология глубоко внедряется в теорию и практику советской медицины, освещая дальнейшие пути развития клинической мысли и обогащая клинические методы исследования и подход к больному новыми, павловскими принципами. Основы этой тесной связи физиологии и клиники были заложены творческим содружеством двух выдающихся русских ученых
И. П. Павлова и С. П. Боткина, объединивших большие творческие коллективы физиологов и клиницистов.
Зарождение физиологического направления в медицине не явилось случайным эпизодом в истории отечественной медицины. Созданию этого направления, его идейному формированию и успешному развитию в значительной мере способствовали передовые идеи замечательных представителей русской науки 60-х годов прошлого столетия Н. Г. Чернышевского, В. Г. Белинского, А. И. Герцена, П. Л. Добролюбова и Д. И. Писарева. Страстная пропаганда естественных наук этими выдающимися прогрессивными деятелями высоко подняла в умах передовой русской интеллигенции того времени роль и значение естествознания как в формировании правильного материалистического миросозерцания, так И в использовании этих знаний для практических целей в жизни народа. На основе этих идей выросли и сформировались такие крупнейшие деятели русской и мировой науки, как Н. И. Пирогов, И. М. Сеченов, П. И. Мечников, Д. И. Менделеев, К. Л. Тимирязев и целая плеяда других выдающихся русских ученых. Эти идеи оказали огромное влияние и на формирование взглядов И. П. Павлова и С. П. Боткина.
Исключительно большое значение в формировании и развитии взглядов Павлова, идеи «нервизма» имели труды выдающегося русского физиолога И. М. Сеченова.
Даже после того как И. П. Павлов стал изучать высшую нервную деятельность, многие его ученики и последователи продолжали исследования пищеварения у животных и человека. Особенно плодотворными явились работы в этой области выдающихся представителей павловской школы Л. А. Орбелп, К. М. Быкова, П. П. Разепкова, Г. В. Фольборта, В. В. Савнча, Г П. Зеленого и многих других.
Трудно перечне, пить все то, чем обогатилась физиология пищеварения за истекшие 60 лет. Для исследования многообразных функций пищеварительной системы разработаны и внедрены новые методические приемы. Творческое использование их позволило глубже проникнуть в интимные механизмы деятельности пищеварительных желёз, гладкой мускулатуры желудочно- кишечного тракта, процессы всасывания и т. д. Значительно полнее стало представление о механизме регуляции деятельности пищеварительной системы и ее взаимоотношений с другими функциональными системами организма.
В изучении процессов, протекающих в пищеварительном аппарате человека и животных, наряду с павловскими классическими методами исследования все большую популярность стали завоевывать другие методы, в частности элсктрофизиологический. Это и попятно, если учесть, что электрические параметры живой клетки бесспорно могут отражать определенные физико-химические сдвиги, происходящие при ее жизнедеятельности. Как известно, переход клетки от состояния относительного покоя к деятельному всегда сопровождается изменением соответствующих величин электрических потенциалов. В. Ю. Чаговец впервые сумел соединить электрофизиологический метод с Павловским приемом изучения функций органов пищеварения. В работах В. Ю. Чаговца и его последователей было установлено, что с помощью метода электрогастрографии (отведения тока от поверхности слизистой оболочки желудка) можно с успехом проследить функциональное состояние желудка. Однако сложность исследования состоит в том, что деятельность секреторных элементов желудка взаимосвязана с его моторными явлениями. И тем не менее в последние десятилетия изучение связи тех и других стало вполне осуществимым. Получены новые экспериментальные факты о характере электрических потенциалов, регистрируемых непосредственно с самих мы печных элементов желудка, и их изменениях при рефлекторном возбуждении деятельности желудочных желез (Bozler, 1962; М. А. Собакин, 1956; П. Г. Богач, 1970; П. К. Климов, 1974, и др.).
Используя электрофизиологический метод, ряду исследователей (В. Е. Делов, В. Н. Черниговский, Л. Д. Ноздрачсв, И. А. Бульнип, Л. В. Итипа, И. А. Алешин и др.) представилась возможность характеризовать афферентные процессы в нервных путях желудка и кишечника, произвести их анализ, установить особенности течения в зависимости от функционального состояния органов брюшной и тазовой полостей.
В последние десятилетия электронно-микроскопическими исследованиями выяснена ультраструктура железистых клеток (К. А. Зуфаров и др.), синтезирующих слизистый, белковый секреты. Изучается синтез, транспорт и формирование мукопротеидных и гликопротеидных секретов.
Оригинальное направление в исследовании деятель-кости органов пищеварения, связанное с менем Г. В. Фольборта, получило дальнейшее развитие в работах Я- П. Склярова с сотрудниками. На примере желудочных желез изучено острое и хроническое перенапряжение секреторного аппарата желудка, а также восстановление работоспособности желудочных желез. Ему принадлежит выяснение особенностей нервных и нервно-химических влияний при измененной работоспособности железистых леток, а также намечены пути ускоренного восстановления работоспособности главных пищеварительных желез.
Ценные сведения получены о динамике желудочной секреции, составе желудочного сока, содержании и распределении в нем основных субстратов, ферментов и ионов. Исследования в этом направлении выполнены Л. Г. Хрипковой с сотрудниками на высоком методическом уровне с использованием методов функциональной гистохимии и других современных биохимических и морфологических методов. Полученные результаты имеют немаловажное значение для понимания механизма образования желудочного сока как в целом, так и отдельных его компонентов.
Обычно секреторная деятельность желудка у человека изучалась в период бодрствования, в последнее время она начала изучаться и в период сна. При этом выяснилось, что во время сна происходит уменьшение объема желудочной и гепато-панкреатико-дуоденальный секреции, связанное развитием в центральной нервной системе сонного торможения, распространяющегося по коре головного мозга и спускающегося на нижележащие центры (Ф. И. Комаров). Результаты этих исследований оказались весьма полезными для клиники.
В настоящее время особенности секреторной функции желудка у людей разного возраста выяснены недостаточно, но тем не менее данные, полученные рядом исследователей (Б. И. Марциповский, Ю. Н. Успенский, Оно Дзенъити, Кренц Фикри и др.), свидетельствуют о том, что у здоровых лиц пожилого возраста при старении происходят функциональные нарушения деятельности пищеварительной системы, в частности снижение кислотности желудочного сока, понижение секреторной реакции желудка как на рефлекторные раздражители, так и гуморально-химические.
Возникающие функциональные изменения со стороны органов пищеварения зависят не только и не только от возраста людей, сколько от возрастных особенностей функционального состояния организма в целом (Б. И. Марциповский). Другие же авторы, не отрицая такую возможность, считают, то в процессе старения происходит вовлечение пищеварительного аппарата и в нем развиваются не только функциональные, но и морфологические изменения (Ю. Н. Успенский и др.).
Работы ряда ученых были посвящены исследованию сезонных колебаний секреции желудочного сока. Эксперименты, проведенные на собаках, выявили зависимость характера желудочной секреции от сезонов года. Так, И. И. Марков обнаружил, что интенсивность сокоотделения у собак увеличивается весной и осенью и снижается летом. Переваривающая сила желудочного сока повышается летом и зимой и понижается весной и осенью. Р. О. Файтельберг, М. М. Стамбольский и Н. И. Гуска более полно проанализировали сезонные сдвиги в работе желудка у собак, исследуя объем секреции желудочного сока, его кислотность, переваривающую силу, содержание в соке астромукопротеидов.
Еще И. П. Павлова интересовал вопрос о зависимости функций органов пищеварения (в частности, поджелудочной железы) от кровоснабжения их. В последующие годы изучению этого вопроса было посвящено немало исследований, позволивших выяснить соотношение секреторной и сосудистой реакций пищеварительных желез при виде и запахе пищи, акте еды, в период разгара пищеварительного процесса и после его окончания, восстановления внепищеварительного уровня. Не остались без внимания и вопросы регуляции кровоснабжения пищеварительных желез.
Исследованиями И.Т.Курцина с сотрудниками получен обширный материал о роли нервной системы в регуляции кровоснабжения слюнных, желудочных и кишечных желез, а также поджелудочной железы. Результаты экспериментального изучения кровоснабжения главных пищеварительных желез имеют определенное клиническое значение, так как позволяют учитывать роль сосудистого фактора в этиологии и патогенезе болезней органов пищеварения.
В одну из наиболее актуальных проблем гастроэнтерологии внесено много новых экспериментальных фактов. В частности, более обстоятельно выяснено долевое участие в кишечном пищеварении внутриполостного и пристеночного или мембранного пищеварения. Внутриполостное пищеварение, его значение и механизмы подвергнуты всестороннему анализу как за рубежом, так и в нашей стране (Г. К. Шльнин, 1967). Что же касается мембранного пищеварения, то, не отрицая роли зарубежных ученых в изучении этого вопроса, тем не менее важно подчеркнуть наиболее крупный вклад в разработку его А.М.Уголева (1967) с сотрудниками и последователями. Он определил не только основные типы пищеварения, но и объяснил, почему они сохранились и развивались в процессе эволюции животного мира. Им показана роль пристеночного пищеварения в деятельности пищеварительного тракта млекопитающих, дана характеристика этого типа пищеварения. Особый интерес представляют сведения о физико-химических и структурных аспектах пристеночного пищеварения. Работы А.М.Уголева получили широкую известность и признание как в нашей стране, так и далеко за ее пределами.
Пищевое поведение человека и животных — одна из сложных общебиологических проблем. Исследованиями последних десятилетий во многом выяснены физиологические механизмы пищевого поведения (П.К. Анохин, А.М. Уголев, П. Г. Богач, А.П.Бакурадзе, А.В.Асатиаш, К.В.Судаков, В.Н.Черниговский, В.Г.Кассиль и др.).
Интенсивно изучается периодическая деятельность пищеварительных органов. В настоящее время дана довольно полная общая характеристика этой деятельности, выяснено функциональное состояние пищеварительного тракта в различные фазы периодического цикла, изменения в других функциональных системах организма, связь периодики с изменениями в обмене веществ.
Значительно продвинулось исследование регуляции периодики, роли периной системы и гуморальных факторов (П.Г.Богач, И.И.Лебедев, К.В.Судаков, И.П.Салмни, А.И.Бакурадзе, А.И.Мордовцев и др.).
В двадцатые годы нашего столетия стали интенсивно разрабатываться вопросы регуляции желчевыделительной системы. В ряде европейских стран развивали теорию нервной регуляции желчевыделительной системы. В этот период в США был открыт специфический гормон холецистокинин и было дано обоснование его участия в регуляции сократительной деятельности желчного пузыря (Айви с соавторами). В 30-е годы в трудах сотрудников К- М. Быкова (А. В. Риккль, С. М. Горшкова, И.Т.Курцин, В. Г. Прокопенко и др.) был накоплен обширный материал по нервной регуляции желчевыделительной системы и зависимости ее функционального состояния от деятельности высших отделов центральной нервной системы. Французскими хирургами получены ценные факты, показывающие значение давления в различных отделах желчевыделительной системы для направления движения желчи. Работы шведских ученых во многом способствовали решению ряда вопросов гормональной регуляции тонуса сфинктера Одди и желчного пузыря. Исследования С. М. Горшковой, Ю. Л. Петровского, Е. Ф. Ларина, А. Н. Бакурад-зе, Н. П. Скакун и других позволили расширить ранее существовавшее представление о секреции желчи, ее физико-химическом составе и свойствах, двигательной функции желчевыделительной системы и механизма ее регуляции.
Следует подчеркнуть практическую важность для клиники работ, выполненных И.Т.Курциным, Л.Д.Лштденбратеном, Л.Я.Губергрицем, И.Б.Шулутко и др.
С большим успехом при изучении механизмов регуляции функций желчевыделительной системы были использованы П. К. Климовым методики рентгенографии и рентгеноскопии, выполненные на современном уровне. Получены новые факты, свидетельствующие о непрерывной работе сфинктера Одди вне пищеварения и прерывистости желчевыделения во время пищеварения. Установлено, что печеночная желчь поступает в желчный пузырь и в период пищеварения. При исследовании механизмов было показано, что на двигательную активность желчного пузыря в процессе наполнения желчевыделительной системы оказываются эфферентные влияния со стороны различных участков коры головного мозга и подкорковых областей мозга. В регуляции принимают участие оба отдела вегетативной нервной системы. С помощью различных фармакологических веществ, а также гормонов получен ряд новых данных о нервно-гуморальном звене регуляции желче-выделительного аппарата.
Роль коры головного мозга в регуляции деятельности пищеварительной системы изучалась в течении длительного времени очень интенсивно (К- М. Быков, В. Н. Черниговский, И.Т.Курцин, М. А. Усиевич и Др.), в результате чего стало особенно понятным ее значение в механизмах приспособительных реакций органов пищеварения к различным внешним и внутренним факторам, выяснению роли важного отдела центральной нервной системы — гипоталамуса — незаслуженно было уделено меньшее внимание. Однако в последние десятилетия интерес к этой важной проблеме значительно возрос. В нашей стране наиболее разносторонние исследования роли гипоталамуса в механизме регуляции пищеварительной системы осуществлены П. Г. Богачем, А. Н. Бакурадзе и др.
Глава 2
Общая характеристика пищеварения и пищевых веществ
Пищеварение — совокупность процессов, обеспечивающих механическое измельчение и химическое расщепление пищевых веществ на компоненты, лишенные видовой специфичности и пригодные к всасыванию и участию в обмене веществ организма животного человека.
Существование человека и животных невозможно без постоянного поступления в организм пищевых веществ, воды, минеральных солей и витаминов. «Сушественнейшей связью животного организма с окружающей природой является связь через известные химические вещества, которые должны постоянно поступать в состав данного организма, т. е. связь через пишу», - говорил И. П. Павлов. Павлов И. П. Поли. собр. соч., т. 111, кн. 1.М., 1951, . 116—117.
Необходимые для жизнедеятельности организма питательные вещества находятся в окружающей природе в виде сложных высокомолекулярных соединений - белков, жиров и углеводов, которые для всасывания и последующего усвоения (ассимиляции) их клетками нуждаются в предварительной механической и химической обработке. Постоянный прием веществ, переработка и усвоение их позволяют сохранить целостность материального субстрата и его жизнедеятельность.
Все живые организмы по типу обмена веществ между организмом и внешней средой разделяются в основ-ком на две группы: автотрофные и гетеротрофные. К первой относятся зеленые растения, которые под влиянием солнечных лучей синтезируют из окружающей неорганической природы и накапливают в себе органические вещества; ко второй — животные, не способные к подобному синтезу и пользующиеся для своего питания уже синтезированными органическими веществами.
В животном организме синтезируются сложные органические вещества из менее ложных, но синтез органических веществ из неорганических у них, за редким исключением, отсутствует, что отличает их от растительных организмов. Однако наряду с этим следует казать на общность многих химических процессов у животных и растений (К. А. Тимирязев).
Химический состав пищевых веществ и их переваривание. Пища, принимаемая человеком, может быть растительного и животного происхождения. В ее состав входят различные питательные вещества: белки, жиры и углеводы, которые, поступая в организм, идут на построение клеток и тканей и служат энергетическими источниками жизнедеятельности организма. Кроме питательных веществ пища содержит воду и неорганические вещества, главным образом в виде солей, которые по являются источником энергии, но без них невозможна нормальная жизнедеятельность клеток и тканей. В ней содержатся добавочные факторы питания — витамины, играющие огромную роль в росте, развитии и функционировании клеток и тканей, а также всего организма в целом.
Составная часть пиши — растительная клетчатка, которая под влиянием ферментов пищеварительных соков не переваривается. Однако это балластное вещество имеет определенное значение в ходе самого пищеварительного процесса, так как оказывает возбуждающее влияние на секрецию и моторику желудочно-кишечного тракта.
Большое участие в превращении пиши в простые растворимые химические соединения принимают биологические катализаторы, или ферменты, которые превращают нерастворимые неспособные к диффузии питательные вещества в растворимые и диффундируюшие соединения. У разных животных этот процесс протекает неодинаково.
Различают внутриклеточное и внеклеточное пищеварение. Внутриклеточное пищеварение свойственно всем одноклеточным и некоторым многоклеточным организмам, находящимся на низких стадиях эволюционного развития. Оно состоит в том, что клетка (или группа клеток) активно захватывает пищевые вещества из окружающей внешней среды и при помощи специальных соков переваривает их, после чего они усваиваются цитоплазмой (рис.
1). В качестве примера можно привести пищеварение у амебы, которая псевдоподиями захватывает пищевые вещества и расщепляет их на простые химические соединения при помощи ферментов, находящихся в клетке. У более дифференцированных одноклеточных, так, например, у парамеции, пищевые вещества из внешней среды поступают в клетку через специальный участок цитоплазмы. В клетке вокруг пиши образуется вакуоль, где и происходит переваривание. Продукты, образовавшиеся в результате этого процесса, частично симиллируется цитоплазмой, а частично вырабатывается наружу в виде отбросов.

Рнс. 1. Внутриклеточное переваривание у Ваlаntіdіm gygan (по Бретшпейдеру и Гиршу):
1-3 — предварительное переваривание пищи; 4-7 — истинное переваривание в вакуолях
Большой интерес представляет внутриклеточное пищеварение у белых кровяных телец (лейкоцитов). В 1887 г. И. И. Мечниковым впервые было показано, что лейкоциты обладают фагоцитарной способностью, т. е. способностью захватывать различные вещества, в том числе бактерии, что имеет важное значение для защиты организма от вреднодействующих агентов внешней среды. У многих беспозвоночных, как, например, у моллюсков и иглокожих, процесс пищеварения протекает в две фазы. Первая фаза происходит внеклеточно, в так называемой пищеварительной полости. Здесь некоторые пищевые вещества (углеводы) размельчаются и расщепляются. Вторая фаза происходит внутриклеточно и состоит, с одной стороны, в ссимиляции уже расщепленных веществ и, с другой,— в переваривании вешесТЁ, не подвергающихся расщеплению в первой фазе. Такой процесс вне- и внутриклеточного пищеварения наблюдается и у некоторых кровососущих насекомых. Во всех этих случаях переваривание пищевых веществ происходит при участии находящихся в цитоплазме особых химических соединений — ферментов.
Внеклеточное пищеварение преобладает у позвоночных и многих беспозвоночных животных и протекает в специальной пищеварительной полости. В этих случаях пищевые вещества, поступая в пищеварительную полость или пищеварительный канал, подвергаются физико-химической обработке, в результате происходит расщепление веществ на простые химические соединения, которые затем всасываются стенками желудочно-кишечного тракта и поступают в жидкости организма (кровь, лимфа), током которых они разносятся всем клеткам тканям.
На низких стадиях развития многоклеточных, как, например, у гидры, пищеварительная полость имеет лишь одно отверстие, через которое и поступает пища и обратно выбрасываются непереваренные остатки. У некоторых червей пищеварительная полость имеет два отверстия: входное, или ротовое, и выходное, или анальное. Между ними находится рубка — кишка, где происходит переваривание пищевых веществ. Вследствие изобилия ферментов на клеточных мембранах пристеночное пищеварение в кишечнике бывает более интенсивным, чем полостное (А.М.Уголев).
Пищеварение у человека и высших животных состоит из трех взаимосвязанных физиологических процессов: 1) секреции пищеварительных соков и воздействия последних на пищевые вещества, 2) моторики желудочно-кишечного тракта и 3) всасывания продуктов пищеварения.
Глава 3
Строение и функции пищеварительного аппарата
По мере развития и совершенствования морфологических структур и функций у животных появляется пищеварительный аппарат, который в зависимости от характера питания и условий существования животного бывает различным. Из переднего отрезка пищеварительной трубки дифференцируются ротовая полость (ротовое отверстие, язык, зубы, жевательные мышцы, слюнные железы и слизистые железы слизистой оболочки полости рта др.), глотка и пищевод. Из средней части развиваются желудок, двенадцатиперстная кишка, поджелудочная железа, печень, тонкая кишка. Задняя часть кишки дифференцируется в слепую кишку, у человека в 5-образную и прямую кишку с анальным отверстием (рис. 2).
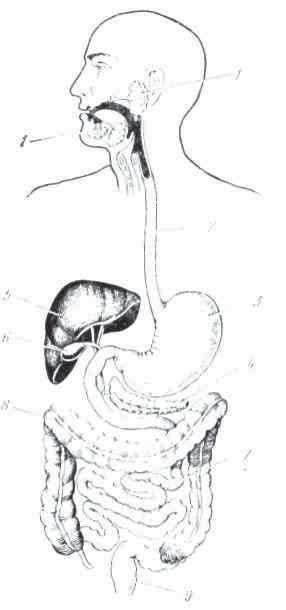
Рис. 2. Схема пищеварительного аппарата у человека: 1 — слюнные железы; 2 — пищевод; 3 — желудок; 4 — поджелудочная железа; 5 — печень; б — желчный пузырь; 7 — тонкие кишки; 8 — толстые кишки; 9 — прямая кишка
На всем протяжении пищеварительной трубки имеются три оболочки: внутренняя — слизистая, средняя — мышечная и наружная — серозная. В переднем ее отделе, оставляющем ротовую полость, имеются, помимо мягких тканей (слизистая, мышечная и р.), хрящевая и костная; в среднем и заднем отделах находятся лишь мягкие ткани (эпителиальная, мышечная, соединительно-тканая и т. п.). Изнутри пищеварительная трубка выстлана слизистой оболочкой, под которой располагается подслизистый слой рыхлой соединительной ткани, где находятся кровеносные сосуды и нервы. Под подслизистым слоем располагается мышечный слой, состоящий на всем протяжении пищеварительной трубки из гладкой мускулатуры, за исключением переднего и заднего концов трубки, где находятся поперечнополосатые мышцы. Мышечная оболочка имеет два слоя: внутренний круговой и на нужный продольный. В желудке три слоя. Серозная оболочка отделяется от мышечной небольшим подсерозным слоем, состоящим из рыхлой соединительной ткани.
Тончайшая дифференциация в развитии пищеварительного аппарата достигается у человека и высших животных тем, что создается морфологическое и функциональное разграничение пищеварительной трубки на специальные отделы, в которых пищевая масса, в зависимости от ее свойств и специализации этих отделов, задерживается на некоторое время ли быстро переводится в следующий. В ряде отделов пищеварительного тракта пища подвергается действию пищеварительных соков, вырабатываемых пищеварительными железами. В каждом отделе имеется своя специализация гладкой мускулатуры, железистых леток, способных выделять в просвет пищеварительной трубки сок.
У человека и высших животных в ротовую полость открываются протоки слюнных желез.
В стенках желудка заложены железы, выделяющие кислый желудочный сок; в верхнем отделе двенадцатиперстной кишки находятся бруннеровы железы, отделяющие щелочной сок; в общей и подвздошной кишках — многочисленные железы, в которых образуется щелочной кишечный сок. Кроме того, в полость двенадцатиперстной кишки открываются протоки поджелудочной железы, выделяющей поджелудочный сок, и печени, выделяющей желчь. В слизистой оболочке по ходу всего желудочно-кишечного тракта расположены железы, выделяющие слизь.
Пищеварительные соки богаты различными ферментами (протеолитические, иполитические и амилолитические), способными расщеплять белковую, жировую и углеводную пищу. Некоторые пищеварительные соки содержат ферменты, действующие только на углеводы (например, слюна), другие, благодаря содержанию нескольких ферментов, обладают способностью действовать на все сорта пищевых веществ, как, например, под- желудочный и кишечный соки, расщепляющие и белки, и углеводы, и жиры.
Наличие в стенке желудочно-кишечного тракта гладких мышц, обладающих способностью сокращаться, обеспечивает механическую обработку пищи и передвижение ее вдоль пищеварительной трубки.
Слизистая тонкого кишечника имеет большое количество складок, углублений (крипт) и ворсинок. Так, в двенадцатиперстной кишке 22—40 ворсинок на 1 мм2.
С помощью электронной микроскопии обнаружено, что каемчатый, или цилиндрический, эпителий, покрывающий поверхность каждой ворсинки, имеет сложное строение. Каемка образована большим числом (1500— 3000 на каждой клетке) цитоплазматичсских отростков— микроворсинок, играющих большую роль в процессе всасывания и переваривания.
Поступившая в пищеварительную трубку пища подвергается действию пищеварительных оков. Секреция пищеварительных соков состоит из связанных между собой процессов: секретообразования и секретовыделения.
По мере эволюционного развития животных наблюдается изменение секреторных клеток, выделяющих сок в пищеварительную полость. У низших животных процессы секреции и всасывания осуществляются эпителиальным слоем средней части кишки, где и происходит пищеварение и всасывание, причем клетки, выполняющие эти две функции, подчас не тдифференцированы друг от друга. Встречается и так, что одна и та же клетка обладает способностью и всасывать продукты расщепления пищи, и выделять пищеварительный секрет. Образование секрета в клетках происходит медленно. Обычно в них накапливаются зерна
(гранулы) или пузырьки жидкости, которые затем выделяются в просвет пищеварительной кишки. Некоторые секреторные клетки при этом погибают. Такой тип секреции, характеризующийся изменением клеточной структуры и нарушением ее целостности, называют морфокинетической секрецией.
Одним из важнейших структурных и функциональных элементов органов пищеварения является секреторная железистая клетка. В каждой пищеварительной железе клетки имеют специфические особенности. Как и и каждой живой клетке, в секреторной непрерывно протекают процессы образования (анаболизма) и распада (катаболизма). В результате ее жизнедеятельности из клетки выделяются неорганические вещества (секреты), участвующие в метаболизме, причем эти вещества (вода, ионы) в цитоплазме не претерпевают химических превращений и выделяются такими же, какими поступают в клетку. В процессе метаболизма в летке, естественно, образуются продукты распада (экскреты), которые по мере накопления должны быть выделены (например, CO2, мочевая кислота, молочная кислота и др.).
А.МУголен отмечает, что наряду с этим в клетке происходит синтез специфических продуктов (секреты), состоящих из более пли менее сложных макромолекул. Образуются они результате анаболических процессов. В некоторых случаях в состав экскретов могут опадать и физиологически активные вещества клетки.
Термин «секреция» подлежит дальнейшему уточнению, тем не менее в настоящее время под секрецией понимается сложный внутриклеточный процесс, в ходе которого секреторная клетка, и в частности пищеварительная, получает из крови исходные вещества, из части которых синтезирует секреторный продукт, выполняющий определенную функцию в организме, и выделяет его вместе с водой и некоторыми электролитами в виде секрета в полость пищеварительной трубки.
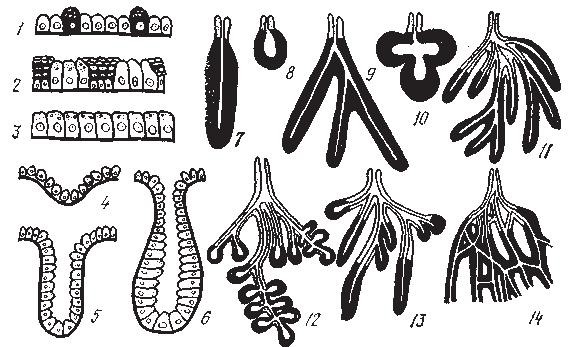
Рис. 3. Пищеварительные железы (по Б. В. Алешину, 1959):
1 — одноклеточные внутриэпителиальные; 2 — железистые почки; 3 — железистое поле; 4 — железистая ямка; 5 — железистая крипта; б — внеэпителиальная многоклеточная рубчатая железа (формы многоклеточных внеэпителиальных желез); 7 — простая трубчатая; 8 — простая альвеолярная; 9 — простая трубчатая с разветвленными концевыми отделами; 10 — простая альвеолярная с разветвленными концевыми отделами; 11— сложная трубчатая; 12—сложная альвеолярная; 13 — сложная трубчатая — альвеолярная; 14 — сложная сетчатая.
Черным заштрихованы ацинусы, светлые — выводные протоки
Существует морфологическая классификация железистых структур пищеварительной системы: одноклеточные, в частности бокаловидные клетки кишечника; многоклеточные, которые подразделяются на: 1) внутриэпителиальные, 2) клеточные поля (секреторный эпителий поверхности желудка), 3) внеэпителиальные многоклеточные железы, малые, концевые отделы которых располагаются в соединительной ткани слизистой оболочки, а выводные протоки открываются на поверхность слизистой (мелкие железы ротовой полости, пищевода, ардиальные, пилорические и фундальные железы желудка, брунне-ровы железы; рис. 3), 4) большие пищеварительные железы, концевые отделы которых располагаются вне слизистой оболочки пищеварительного тракта, а длинные выводные протоки открываются в просвет пищеварительной трубки (слюнные железы, поджелудочная железа, печень).
За последние годы многое стало известно об ультраструктуре клеток, синтезирующих секрет разного состава (белковый, слизистый). Секреторные клетки имеют весьма сложную структуру, причем в секреторном процессе участвуют все органоиды (рис. 4). В структуре и обменных процессах клеток происходят закономерно повторяющиеся изменения, связанные с образованием и выделением секрета. Комплекс этих изменений получил название секреторного цикла. Следует, однако, отметить, что до настоящего времени в литературе нет единой точки зрения о фазах или периодах секреторного цикла в ритмически функционирующих железистых клетках. Различают два типа секреции: непрерывный и прерывистый. При непрерывном типе (секреторные клетки поверхностного эпителия желудка) секрет выделяется по мере синтеза; при этом одновременно наблюдаются все фазы цикла — поглощение материала, внутриклеточный синтез и выделение секрета. При прерывистом или ритмическом типе (например, бокаловидные слизистые клетки кишок) цикл растянут во времени и происходит в определенной последовательности. Синтез новой порции секрета начинается после выведения предыдущей. Большинство клеток мерокриновых и покриновых желез после выделения запасов секрета вновь приступают к его синтезу. Однако клетках с различным типом секреции восстановительные процессы протекают неодинаково.
В мерокриновых железах восстановление идет параллельно выработке секрета, а в покриновых оно происходит после прекращения секретообразования.
В настоящее время имеются данные, позволяющие судить о продолжительности различных фаз секреторного цикла, они могут продолжаться в течение нескольких часов.
Пищеварительные железы обильно снабжены кровеносными сосудами, из которых в секреторные клетки ацинусов поступают вода, неорганические вещества и бизкомолскулярпые органические соединения — аминокислоты, моносахариды, жирные кислоты. Они проходят через ряд барьеров, располагающихся между просветом капилляров и цитоплазмой секреторной клетки. В процессе транспорт та веществ из капилляра наблюдается азрыхление его базальпоп мембраны, расширение субэндотелиальных клеток с образованием енестр, значительное увеличение пиноцитозпых пузырьков. Изменяется и структура эндотелиальных клеток. Далее вещества проходят периканиллярное пространство, базальную мембрану линчует и базальную плазменную мембрану секреторной клетки. Назальная мембрана капилляров и ацинусов, а также плазматическая мембрана секреторной клетки выполняют важную функцию проницаемости. По-видимому, транспорт вещества через мембрану является активным процессом, требующим энергетических затрат. Не случайно в иноцитозных пузырьках эндотелиальных клеток высокая активность АТФазы. Поступление веществ в железистую клетку, вероятно, осуществляется как с помощью пиноцитоза, так и путем диффузии. При пиноцитозе поступают крупномолекулярные соединения вещества; диффузия обеспечивает прохождение более мелких молекул. В переносе вещества участвует елочная фосфатаза, особенно в период его активного поступления в клетку.
Т.А. Григорьева выделяет следующие морфологические признаки, характеризующие< процесс поступления вещества в железистую клетку: 1) присутствие пиноцитозных пузырьков в цитоплазме железистой клетки вблизи ее сосудистого полюса, 2) наличие в той или иной степени впячиваний плазмолеммы базалыной части железистой клетки, 3) изменение активности АТФазы и щелочной фосфатазы в клеточных мембранах. Поступающие в железистую клетку вещества — исходный материал не только для образования секреторного продукта, но и для обеспечения обмена в железистой клетке.
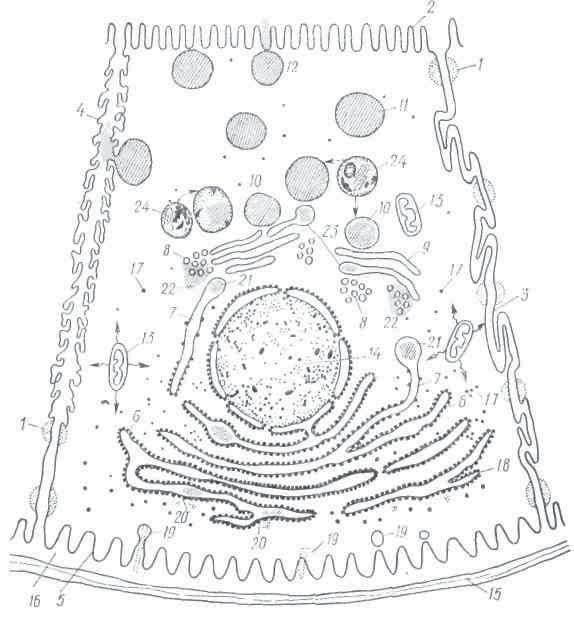
Рис. 4. Экзокринная секреторная клетка пищеварительной железы, синтезирующая белковый субстрат (по Е. А. Шубппковой, 1967):
1 — десмосомы; 2 — микроворсинки; 3 — боковые кладки плазмолеммы; 4 — секреторный капилляр; 5 — пячивания базалыюй лазмолеммы; б — гранулярный ндоплазматическнй рстпкулум; 7— промежуточный элемент эидо-плазматического ретикулума; 8 — пузырьки аппарата Гольджи; 9 — мембраны аппарата Гольджи; 10 — конденсирующие вакуоли аппарата Гольджи; 11 — секреторные гранулы; 12 — выход секрета в секреторные капилляры и полость концевого отдела; 13 — митохондрии; 14 — ядро; 15 — базальная мембрана; 16 — цементирующие вещества; 17 — свободные рибосомы; 1$ — рибосомы, связанные с эпдоплазматическим етикулумом; 19 — исходные вещества, поступающие в клетку для синтеза; 20 — начало синтеза белка па рибосомах гранулярного ретикулума и проникновение предшественника секрета внутрь его канальцев и цистерн (предшественник и секреторный продукт изображены штрихами); 21 — перемещение предшественника белкового секрета в промежуточный элемент; 22 — появление предшественника секрета в области мелких пузырьков Гольджи, 23 — накопление секреторного продукта в расширениях мембран и вакуолях Гольджи; 24 — изосомы (мелкие точки — гидролазы). Прерывистая стрелка — перемещение гидролаз из изосом в незрелые секреторные гранулы или вакуоли Гольджи; зигзагообразные стрелки — выход энергии из митохондрий в настоящее время изучены процессы синтеза ОСЛКОВОГО секрета в поджелудочной железе околоушной слюнной железе. Изменения в секреторных клетках начинаются в ядрышке дер, которые увеличиваются в размерах и начинают давать интенсивную реакцию на РНК.
Эта рибосомпая РНК (рРНК) соединяется с белком и в виде рибосом поступает в цитоплазму.
На молекулах ДНК ядра идет синтез информационной РНК (ИРПК) и транспортной РНК
(тРНК) рибонуклеиновых кислот, которые несут генетическую информацию о первичной структуре белка к синтетическому аппарату клетки— свободным рибосомам и рибосомам гранулярного эндоплазматического ретикулума. К рибосомам подходит иРПК, объединяет их комплексы — полисомы, на которых осуществляется синтез структурных белков из аминокислот.
В ацинарных клетках осуществляется синтез двух видов белков— структурного, используемого для построения компонентов клетки, и секреторного, выделяющегося из летки в виде гранул секрета.
В образовании секрета принимает участие ядро клетки, во-первых, в ядре образуется первичный секреторный продукт (просекрет), окончательное формирование которого осуществляется в цитоплазме; во-вторых, в нем, как в центре генетической информации клетки, синтезируются вещества, без которых невозможно синтезировать секрет в цитоплазме.
Синтез, транспорт и формирование слизистого секрета (мукополисахариды, ликопротеиды) в клетках пищеварительных желез происходит в пластическом комплексе
Гольджи. Предполагают, что многие полисахариды синтезируются в различных структурных компонентах клетки. Однако углеводный компонент гликопротеидов клеток печени, как и мукополисахаридов, образуется в комплексе Гольджи.
Фаза накопления секрета. В железах с циклической секрецией гранулы или вакуоли концентрируются в ацинарных клетках и секрет по мере необходимости выделяется в просвет протоков или в полости пищеварительного тракта. Ацинарные клетки выполняют роль ременного депо пищеварительных ферментов, так как в железе, как правило, отсутствуют специальные участки для хранения секреторных гранул.
Фаза выделения (экструзии) секрета из железистой клетки. Существует классификация желез в зависимости от типа выделения ими секрета. В частности, Ранвье разделил их на два типа: голокриновые и мерокриновые. В дальнейшем мерокриновые железы были подразделены на два типа: апокриновые и собственно мерокриновые.
Применение электронной микроскопии и авторадиографии позволило Куросуми в зависимости от механизма выделения секрета из клетки различать пять типов секреции: 1. Голокриновый — вся клетка превращается в секрет в результате ее дегенерации. Этот тип секреции характерен для поверхностного эпителия желудка. 2. Апокриновый — выделение вместе с секретом части цитоплазмы клеток. Такой тип секретов характерен для протоков слюнных желёз человека в эмбриогенезе. 3. Мерокриновый — выделение секрета не сопровождается разрушением клетки или отрывом части цитоплазмы. После выхода секрета, накопившегося в гранулах и вакуолях, клетка снова приступает к его выработке. Этот тип секреции характерен для большинства пищеварительных желёз. Четвёртый и пятый типы — с разным выходом секрета (рис. 5).
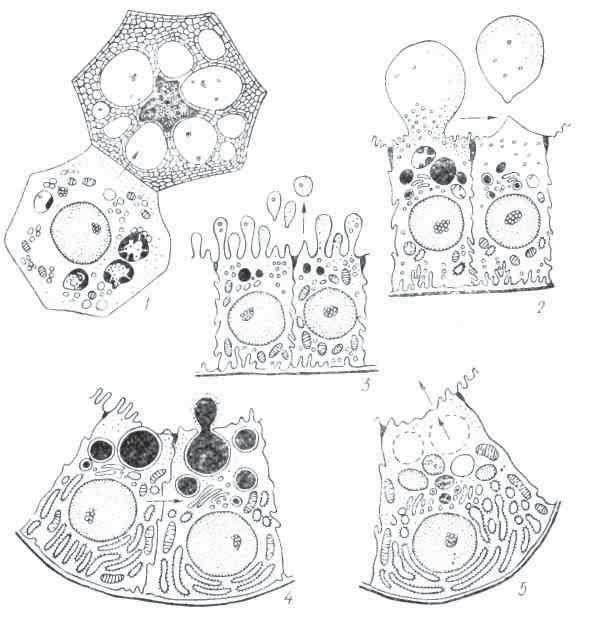
Рис. 5. Типы выведения секрета (по Киковиті, 1961):
1 — голокриновый; 2 — макроапокриновый; 3 — микроапокриновый; 4 — мерокриновый с выходом секрета через отверстие в клеточной мембране; 5 — мерокриновый с выходом секрета через неповрежденную плазматическую мембрану. Стрелка — путь выведения секрета
Большинство клеток мерокриновых и апокриновых желез после выделения запасов секрета вновь приступает к его синтезу. Поэтому можно говорить об их циклической деятельности. В клетках с различным типом секреции восстановительные процессы протекают неодинаково. Как указывалось, в мерокриновых железах восстановление идет параллельно выработке секрета, а в апокриновых оно происходит после прекращения секретообразования (Е. А. Шубникова).
Электрофизиологическими исследованиями, осуществляемыми с помощью микроэлектродной методике, установленного, что мембранный потенциал секреторных клеток различных желез пищеварительного тракта варьирует 'в широких пределах — от 10 до 80 мВ.
В большинстве невозбужденных железистых клеток поляризация не превышала 30—35 В. Пока не получили объяснения низкой величины мембранного потенциала большинства железистых клеток. Вероятно, они связаны со спецификой выполняемой ими функции — с непрерывными процессами восстановления клеточной структуры и пополнением секретированных ею компонентов.
В состав пищеварительных соков входят вода, органические вещества и соли. Из органических веществ наиболее существенное значение имеют ферменты, которые относятся к группе гидролаз, т. е. ферментов, способных присоединять к своей структуре Н+ и ОН-, превращая таким образом нерастворимые вещества в растворимые. Ферменты являются катализаторами биохимических процессов. В зависимости от действия на те или иные вещества они делятся на: амилолитические, или амилазы, расщепляющие углеводы; ротсолитические, или протеазы, расщепляющие белки, протеины, и липолитические, или липазы, расщепляющие жиры, липиды.
Основные, свойства ферментов следующие: ускорение биохимических процессов; специфичность действия, т. с. расщепление или синтез только определенных веществ; стойкость в условиях высокой температуры; активность в определенной среде (кислой, елочной, нейтральной).
Большинство ферментов является комплексными соединениями, состоящими из так называемой группы и компонента, имеющего белковую природу. Их активность зависит от наличия и сохранности в структуре белковой группы сульфгидрильных групп. Характер действия ферментов обусловливается температурой среды, ионным составом сока. В частности, с повышением температуры до определенного предела интенсивность действия их повышается.
Оптимумом действия ферментов является температура от 35 до 50° С. Пищеварительные ферменты холоднокровных и теплокровных животных не идентичны.
Ряд ферментов выделяется в неактивном состоянии и для проявления своего действия требует наличия активаторов. Неактивное состояние пищеварительных ферментов в секреторных клетках, по-видимому, обусловливает устойчивость самих желез к переваривающему действию ферментов. Возможно, известную роль играет и наличие в тканях особых веществ — парализаторов, тормозящих действие ферментов. По данным некоторых авторов, в слизистой желудка содержатся антиферменты, предохраняющие ее от переваривания ферментами желудочного сока.
У простейших расщепление белков, жиров и углеводов различными ферментами происходит в пищеварительной вакуоли, причём смена действий ферментов обуславливается заменой реакции в самой вакуоли. У высших животных этот процесс расщепления пищевых веществ осуществляется в различно дифференцированных отделах пищеварительной трубки.
Таким образом, развитие функции и дифференциация клеток и тканей организма в связи с непрерывно меняющимися условиями внешней среды обусловливают появление качественно новых физиологических процессов, причем между структурой и физиологическим процессом, т. е. между формой и функцией, существует тесная взаимосвязь и взаимообусловленность.
Строгая специализация различных пищеварительных ферментов и сложность пищеварительного процесса у животных в связи со своеобразным типом их питания являются примером приспособительной эволюции функций.
Глава 4
Пищеварение в ротовой полости и глотание
Передний отдел пищеварительного аппарата — ротовая полость — является начальным отделом пищеварительной трубки, куда при естественных условиях прежде всего поступает пища и где она подвергается первоначальной механической и химической обработке. Поступившие в рот горькие, соленые, кислые и сладкие вещества производят раздражение вкусовых рецепторов (окончаний чувствительных нервов), расположенных в различных зонах слизистой языка (рис. 6). Кроме того, в слизистой ротовой полости имеются нервные окончания, воспринимающие температурные и механические раздражения. Возникающие при раздражении рецепторов слизистой рта нервные импульсы передаются по центростремительным, афферентным нервам (тройничному, лицевому и языкоглоточному) в центральную нервную систему, вплоть до коры больших полушарий головного мозга, где возникает ощущение того или иного вкусового качества поступившего в рот вещества (горького, соленого, сладкого или кислого). Из центральной нервной системы импульсы по центробежным, эфферентным нервам направляются к мышцам и слюнным железам и возникают жевательные, сосательные движения и выделение слюны. Весь этот процесс является сложным рефлекторным актом. В результате ощущения, возникшего в связи с тем или иным вкусовым качеством веществ, последние или выбрасываются изо рта наружу — отвергаемые (непищевые) вещества, или подвергаются механической и химической обработке — пищевые (съедобные) вещества.
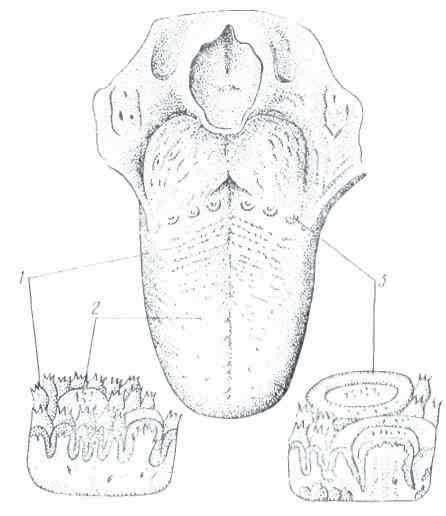
Рис. 6. Сосочки языка: 1 — нитевидные; 2 — грибовидные; 3 — желобоватые
Пища находится в ротовой полости сравнительно короткое время (10—25 с), тем не менее достаточное для того, чтобы она могла быть размельчена и смочена слюной, для формирования пищевого комка, т. е. подготовлена к проглатыванию. Пищеварение во рту сводится главным образом к механической обработке пищи. Химическое же воздействие слюны на пищевые вещества (углеводы) весьма ничтожно из-за непродолжительного пребывания пищевой массы в ротовой полости. Однако переваривающее действие слюны, поступившей вместе с пищевым комком в желудок, продолжается еще в течение некоторого времени, т. е. до тех пор, пока не наступит кислая реакция.
Следует подчеркнуть, что хотя химическое расщепление веществ во рту и весьма незначительное, однако и все связанное с приемом пищи — вид, запах, обстановка, звуки, жевание, раздражение вкусовых рецепторов языка, механических и термических рецепторов слизистой полости рта, глотки и др.— имеет большое значение для последующего хода пищеварительного процесса, так как акт еды — мощный рефлекторный возбудитель деятельности секреторных клеток желудка, поджелудочной железы, тонкой кишки и печени, а также гладкой мускулатуры пищеварительного тракта.
Жевание — сложный рефлекторный акт, состоящий из последовательных сокращений жевательной мускулатуры. Движение нижней челюсти происходит не только по вертикали, но и по горизонтали, в результате чего пища разрывается на части и тщательно перетирается зубами обеих челюстей.
Жевание имеет важное значение для последующего хода пищеварения. Оно способствует оценке вкусовых качеств пищи, стимулирует отделение пищеварительных соков, которые быстрее и лучше проникают в размельченную пищу, а это обусловливает более полное переваривание и последующее ее всасывание. Хорошо механически обработанная и пропитанная слюной пищевая масса затем проглатывается. И. П. Павлов придавал большое значение акту еды.( Неторопливая еда и тщательное пережевывание пищевой массы в ротовой полости способствуют дальнейшему пищеварению и предохраняют слизистую желудка от заболеваний (например, гастрита).
Нервный центр жевания находится в продолговатом мозге. Однако способность произвольно регулировать функцию жевания, сознательно влиять на нее позволяет предположить, что представительства для акта жевания имеются в структурах различных уровней мозга, в том числе и коре головного мозга.
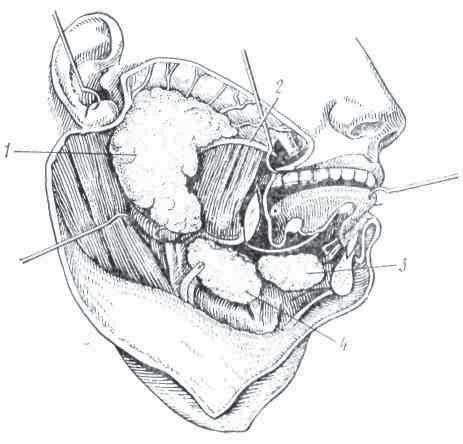
Рис. 7. Слюнные железы человека: 1 — околоушная; 2 — проток околоушной железы; 3 — подъязычная; 4 — подчелюстная.
Процесс сосания так же, как и процесс жевания, — рефлекторный акт. Он имеет особенно большое значение у детей грудного возраста. У взрослых он имеет значение лишь при приеме жидкой пищи. В акте сосания принимает участие мускулатура рта и языка, которая при сокращении создает в ротовой полости разрежение воздуха (до 100 — 150 мм вод. ст.).
Слюнные железы. У человека имеются три пары крупных слюнных желез: околоушные, подъязычные и подчелюстные (рис. 7). У собак и хищных животных имеются, кроме того, орбитальные или глазничные, протоки которых открываются в полость рта.
Слюнные железы состоят из слизистых и серозных клеток. Первые выделяют секрет густой консистенции, вторые — жидкую серозную, или белковую, слюну. Околоушная железа содержит только серозные клетки. Такие клетки находятся и на боковых частях языка.
Подчелюстная и подъязычная железы являются смешанными железами, содержащими как серозные, так и слизистые клетки. Смешанные железы располагаются также и в слизистой оболочке губ, щек и кончика языка.
Физиологическое значение слюнных желез прежде всего определяется участием в процессе пищеварения (секреторная функция). Кроме того, они способны выделять из организма некоторые продукты обмена веществ (выделительная функция), а также вырабатывать и выделять в кровь специальный гормон, стимулирующий углеводный обмен в организме (инкреторная функция). У некоторых животных, например у собак, не имеющих на поверхности тела потовых желез, слюнные железы участвуют в процессе терморегуляции организма (терморегуляторная функция).
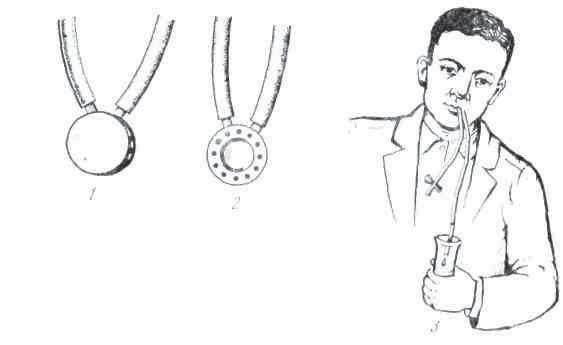
Рис. 8. Получение слюны у человека через капсулу П. И. Красногорского: 1 — вид капсулы сверху; 2 — снизу; 3 — собирание слюны
Методика изучения слюноотделения у животных и человека
У животных слюна может быть получена в остром и хроническом опыте. В первом случае слюна добывается путем вставления в слюнной проток стеклянной канюли, во втором — утем наложения постоянной слюнной фистулы по способу, разработанному в лаборатории И. П. Павлова Д. Л. Глинским в 1895 г.
Операция наложения постоянной фистулы протока слюнной железы, например околоушной, производится следующим образом. У собаки вырезается участок слизистой оболочки рта диаметром 2-3 см с местом впадения слюнного протока; затем последний от- репаровывается от окружающих тканей и выводится в кожную рапу на наружную поверхность щеки, где и фиксируется швами. Слюна, отделяющаяся наружу, может быть получена в изолированном и чистом виде для количественного и качественного анализа.
Эта методика широко применяется в физиологических лабораториях при изучении словных рефлексов у собак. К ней прибегают и при изучении слюноотделения у сельскохозяйственньтх животных.
Хронические фистулы протока слюнных желез иногда встречаются и у человека после ранений или при воспалительных процессах. Обычно же исследование слюноотделения у человека производится при помощи маленькой металлической воронки, или капсулы Н. И. Красногорского, которая накладывается на участок слизистой оболочки, где открывается проток слюнной железы (рис. 8). Капсула имеет форму диска диаметром в 18 мм и состоит из двух камер: внутренней, находящейся против отверстия протока, и наружной, которая при помощи резиновой трубки сообщается с сосудом, где разрежается воздух и создаются условия ля присасывания капсулы к слизистой. Отделяющаяся слюна попадает во внутреннюю камеру, а оттуда по резиновой трубке — наружу в градуированный цилиндр. Присутствие такой капсулы во рту не препятствует приёму небольших количеств жидкости и плотной пищи, что удобно при исследовании действий разных веществ, находящихся в ротовой полости, на слюноотделении.
Состав и свойства слюны
Слюна бесцветная, слегка опалесцирующая жидкость щелочной реакции (pH = 7,4 8,0), не имеющая запаха и вкуса. Она может быть густой, вязкой, подобно слизи, или, наоборот, жидкой, водянистой. Консистенция слюны зависит от неодинакового содержания в ней белковых веществ, главным образом гликопротеида муцина, который придаёт слюне слизистые свойства. Муцин, обволакивая и пропитывая пищевой комок, обеспечивает его свободное проглатывание. Кроме муцина в состав слюны входят неорганические вещества - хлориды, фосфаты, карбонаты натрия, калия, магния и кальция, азотистые соли, аммиак и органические - глобулин, аминокислоты, креатинин, мочевая кислота, мочевина и ферменты.
Плотный остаток слюны равен 0,5-1,5%. Количество воды колеблется от 98,5 до 99,5%. Плотность равна 1,002-0,008. В ней находится некоторое количество газов: кислород, азот и углекислота. У человека и некоторых животных в состав слюны входят еще роданистый калий и натрий (0,01 %). В состав слюны входят ферменты, под влиянием которых перевариваются некоторые углеводы. В слюне человека имеется амилолитический фермент птиалин (амилаза, диастаза), который гидролизует крахмал, превращая его в декстрины и дисахарид - мальтозу, которая под действием фермента мальтазы расщепляется до глюкозы. Расщепление варёного крахмала идет энергичнее, чем сырого. Птиалин действует на крахмал в щелочной, нейтральной и слабокислой среде. Оптимум его действия находится в пределах нейтральной реакции.
Образование фермента происходит главным образом в околоушных и подчелюстных железах.
Хлористый натрий усиливает, а слабые концентрации соляной кислоты (0,01%) ослабляют переваривающее действие фермента. При наличии высоких концентраций соляной кислоты фермент разрушается, поэтому, попадая в желудок, в желудочном соке которого высокая концентрация соляной кислоты (0,5%), слюна вскоре теряет свои ферментативные свойства.
Кроме птиалина и мальтазы в слюне человека содержатся протеолитический и липолитический ферменты, действующие соответственно на белковую и жирную пищу.
Однако практически их переваривающее действие весьма слабо.
В слюне содержится фермент лизоцим, обладающий бактерицидным действием. По представлению И. П. Павлова, слюна обладает лечебным действием (с этим, по-видимому, связано зализывание ран животными).
В процессе секреции слюны обычно различают два момента: перенос воды и некоторых электролитов крови через секреторные клетки в просвет железы и поступление органического материала, образованного секреторными клетками. Известны прямое влияние ионной концентрации солей в крови на состав слюны, нервная регуляция концентрации слюны, обусловленная активностью мозговых центров, регулирующих содержание солей в крови, и, наконец, действие минералокортикоидов на концентрацию солей в крови. Под влиянием кортикоидов надпочечных желез может повышаться в слюне концентрация калия и понижаться концентрация натрия. Под влиянием нервного раздражения или гуморального воздействия клетки слюнных желез могут становиться проницаемыми для неэлектролитов, в частности для некоторых веществ (белков) с высокой молекулярной массой. При попадании в рот отвергаемых веществ слюна нейтрализует их, разбавляет и смывает со слизистой рта — в том заключается большой биологический смысл слюноотделения.
Общее количество выделяемой за сутки слюны у человека составляет приблизительно 1,5 , а у крупных сельскохозяйственньтх животных от 40-60 до 120 л.
Возбудители секреции слюны.
Секреция слюны начинается через 1-3 с после начала действия раздражителя. Это время в физиологии называют скрытым или латентным периодом. По сравнению с другими пищеварительными железами слюнные имеют самый короткий латентный период секреции. По данным некоторых авторов, он исчисляется даже долями секунды. Это свидетельствует о высокой реактивной способности этих желёз. Состав слюны может быть различным в зависимости от физических и химических свойств веществ, поступающих в ротовую полость (С. Г. Вульфсон, Н. М. Гейман, А. П. Зельгейм, Д. С. Фурсиков). Наибольшее количество слюны выделяется на сухие сорта пищи. Поэтому на сухари обычно слюны бывает больше, чем на мягкий хлеб. Точно так же мясо вызывает меньшее отделение слюны, чем мясной порошок. Значительные количества жидкой водянистой слюны выделяются на такие отвергаемые вещества, как соляная кислота, щелочь, песок (табл. 2).
Зависимость секреции от сухости пищевых веществ была установлена и у сельскохозяйственных животных с постоянной фистулой протока околоушной железы.
На примерах, приведенных в табл. 2, можно убедиться в существовании определенной приспособляемости работы слюнных желез к сорту пищи. Этот важный факт был установлен И. П. Павловым не только в отношении слюнных, но и других пищеварительных желез.
У людей слюна выделяется непрерывно (0,1- 0,2 мл/мин). При поступлении веществ в ротовую полость количество отделяемой слюны увеличивается, причем сухость пищи не играет такой роли, как у животных, что, по-видимому, связано с кулинарной обработкой пищи. Значительное отделение слюны происходит лишь на кислые вещества. Вода вызывает отделение вязкой слюны. Жевание усиливает слюноотделение, причем чем больше измельчается пища, тем значительнее отделение слюны.
Умственная и физическая работа уменьшает слюноотделение. Количество и качество отделяемой слюны у человека также находится в зависимости от сорта пищи (табл. 3).
Итак, величина слюноотделения и состав слюны зависят от физико-химических свойств пищи. Кроме того, интенсивность секреции находится в зависимости от общего состояния организма. Так, например, если собаку долго держать на углеводном режиме питания, то в ее слюне появляется птиалин, который обычно отсутствует (И. П. Разенков). При ограничении питья у собак резко повышается реакция слюнных желез на действие воды (Д. С. Фурсиков).
Слюноотделение — рефлекторный акт, который осуществляется при помощи центральной нервной системы, центростремительных (афферентных) и центробежных (эфферентных) нервов. Под влиянием механических, химических и термических раздражений слизистой полости рта в нервных окончаниях (рецепторах) слизистой возникают импульсы, которые направляются по афферентным нервам в центры слюноотделения, откуда возвращаются к слюнным железам по эфферентным нервам.
Разнообразные раздражители неодинаково воспринимаются рецепторами различных участков слизистой оболочки рта. Слизистая языка и часть поверхности слизистой оболочки рта обладают большей возбудимостью к химическим раздражителям. Горькие и солёные вещества вызывают слюноотделение главным образом с корня языка. Терморецепторы находятся почти на всей поверхности языка; Механорецепторами обильно снабжены слизистая корня и кончика языка, мягкое и твёрдое нёбо.
Таблица 3. Слюноотделение (в течение 5 мин) у человека при еде или введении в полость рта различных веществ(по А. Д. Бирюкову)
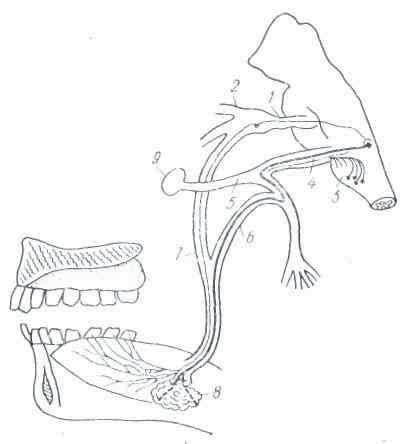
Рис. 9. Рефлекторный нервный путь слюноотделения подчелюстной слюнной железы (по Мюллер, 1924): 1 — тройничный нерв; 2 — Гассеров зел; 3 — ядро лицевого нерва; 4 — лицевой нерв; 5 — коленчатый узел; б — барабанная струна; 7 — язычный нерв; 8 — слюнная подчелюстная железа и подчелюстной слюнной ганглий; 9 — крыло-нёбный ганглий. Тонкая линия — чувствительные нервные волокна, идущий от рецепторных аппаратов языка к чувствительной клетке, расположенной в Гассеровом узле; толстая линия — преганглионарное пересимпатическое волокно, идущее к нервным клеткам в пересимпатическом подчелюстном узле; прерывистая линия — постганлионарные пересимпатические волокна в подчелюстной е тезе и верхняя гу6а Афферентными нервами слюнных желез являются язычный (ветвь тройничного) и языкоглоточные нервы, а также верхняя гортанная ветвь блуждающих нервов и барабанная струна. Кроме того, раздражение других чувствительных нервов может рефлекторным путём вызывать слюноотделение. Эфферентными нервами слюнных желёз являются парасимпатические и симпатические нервы (рис. 9, 10).
В отношении парасимпатической иннервации слюнных желез известно следующее: нейроны, от которых отходят преганглионарные волокна, снабжающие смешанные железы, расположены в верхнем слюноотделительном ядре продолговатого мозга. В составе барабанной струны эти волокна достигают подъязычного и подчелюстного узлов, расположенных в теле каждой из одноименных желез.
В этих ганглиях находятся нейроны, от которых отходят постганглионарные нервные волокна, оканчивающиеся на клетках слюнных желез. В нижнем слюноотделительном ядре продолговатого мозга расположены нейроны, от которых отходят преганглионарные волокна околоушной железе. В составе языкоглоточного нерва и его веточки — малого поверхностного каменистого нерва эти волокна доходят до ушного узла. В этом ганглии расположены нейроны, от которых отходят постганглионарные волокна. В составе височно- ушного нерва эти волокна достигают околоушной железы.
Симпатическая иннервация слюнных желез состоит в следующем: нейроны, от которых отходят преганглионарные волокна, расположены в боковых рогах спинного мозга на уровне ThII-TVI. Волокна подходят к верхнему ганглию, где заканчиваются па постганглионарных нейронах, дающих начало аксонам. Вместе с сосудистым сплетением, сопровождающем внутреннюю сонную артерию, волокна достигают околоушной слюнной железы в составе сосудистого сплетения, облегающего наружную сонную артерию, подчелюстной и подъязычной слюнных желез.
Раздражение черепномозговых нервов, в частности барабанной струны, вызывает значительное выделение жидкой слюны. Раздражение же симпатических нервов вызывает незначительное отделение густой слюны с обильным содержанием органических веществ.
Нервные волокна, при раздражении которых происходит выделение воды и солей, называются секреторными, а нервные волокна, при раздражении которых выделяются органические вещества,— трофическими. При длительном раздражении симпатического или парасимпатического нерва происходит обеднение слюны органическими веществами.
Таблица 4. Дыхание слюнной железы в покое и деятельном состоянии
| Состояние железы | Поглощение кислорода а 1 мин, мл | Выделение Углекислоты за 1 мин, мл |
| Покой | 0,25 | 0,27 |
| Деятельность | 0,86 | 0,97 |
Если предварительно производить раздражение симпатического нерва, то последующее раздражение парасимпатического нерва вызывает отделение слюны, богатой плотными составными частями. Это же происходит и при одновременном раздражении обоих нервов. На этих примерах можно убедиться в той взаимосвязи и взаимообусловленности, которые существуют при нормальных физиологических условиях между симпатическими и парасимпатическими нервами в регуляции секреторного процесса слюнных желез.
При перерезке секреторных нервов у животных наблюдается через сутки непрерывное, паралитическое отделение слюны, которое продолжается около пяти-шести недель. Это явление, по-видимому, связано с изменением в периферических концах нервов или в самой железистой ткани. Возможно, что паралитическая секреция обусловлена действием циркулирующих в крови химических раздражителей. Вопрос о природе паралитической секреции требует дальнейшего экспериментального изучения.
Слюноотделение, возникающее при раздражении нервов, является не простой фильтрацией жидкости из кровеносных сосудов через железы, а сложным физиологическим процессом, возникающим в результате активной деятельности секреторных клеток и центральной нервной системы. Доказательством этого является тот факт, что раздраженно нервов вызывает слюноотделение и после того, как полностью перевязываются сосуды, снабжающие кровью слюнные железы. Кроме того, в опытах с раздражением барабанной струны было доказано, что секреторное давление в протоке железы может быть почти в два раза большим, чем кровяное давление в сосудах железы, однако секреция слюны в этих случаях бывает обильной.
При работе железы резко увеличивается поглощение кислорода и выделение углекислоты секреторными клетками (табл. 4). Количество протекающей через железу крови во время деятельности увеличивается в 3—4 раза.
Микроскопически было установлено, что в период покоя в железистых клетках накапливаются значительные количества зерен секрета (гранулы), которые во время работы железы растворяются и выделяются из клетки.
Секрстообразование и секретовыделение— взаимосвязанные процессы. В опытах показано, что если раздражать, например, барабанную струну, то наряду с интенсивной секрецией слюны наблюдается резкое изменение электрического потенциала слюнной железы. Однако подобное изменение электрического потенциала происходит вскоре же после прекращения выделения секрета, как результат протекающих в клетках железы химических процессов, связанных с накоплением секрета, с секретообразовательной функцией железы.
При раздражении секреторных нервов слюнных желез в последних образуются биологически активные вещества (медиаторы) типа ацетилхолина‚ которые при введении в ток крови, идущей к железе, могут вызвать секрецию, подобную секреции, возникающей при нервной стимуляции. Медиаторы являются химическими передатчиками возбуждения с нерва на железистую ткань. Они образуются при возбуждении или торможении не только периферических нервов, но и центральной нервной системы. Помимо ацетилхолиноподобных веществ, образующихся обычно при раздражении парасимпатических нервов, могут образовываться адреналиноподобные вещества при раздражении симпатических нервов.
В регуляции слюноотделения, осуществлении безусловных и условных слюноотделительных рефлексов важнейшая роль принадлежит центральной нервной системе, в особенности ее высшему отделу — коре больших полушарий головного мозга.
В регуляции слюноотделения участвует диэнцефаль-ная область. Так, например, при стимуляции переднего отдела гипоталамуса или преоптической области стимулируется механизм теплопотерь, о чем свидетельствует появление у животного одышки, слюноотделения при открытой пасти. При стимуляции заднего отдела гипоталамуса на фоне эмоционального возбуждения наступает увеличение слюноотделения.
Стимуляция в условиях хронических опытов на собаках участков гипоталамуса, окружающих воронку, вызывала слюноотделение из ипсилатеральных слюнных желез (П. Г. Богач, А. Ф. Косенко). При стимуляции амигдалярного комплекса наблюдалось в числе других пищевых реакций и слюноотделительная.
Исследованиями И. П. Павлова и других было показано, что раздражение различных участков коры головного мозга (около сильвиевой борозды, верхней фронтальной извилины, орбитальной извилины и лобной доли) сопровождалось отделением слюны. П. Г. Богач и А. Ф. Косенко выявили возможность тормозящих влияний со стороны мозговой коры на гипоталамические отделы слюноотделительного центра (после удаления лобных долей коры раздражение латерального гипоталамуса обусловливало усиление слюноотделительного эффекта).
Возбуждение центров слюноотделения наступает в естественных условиях как при поступлении центростремительных импульсов по нервам, так и при изменениях химического состава крови, т. е. гуморальным путем. Установлено, например, что углекислота, накапливающаяся в крови во время удушья, возбуждает центры слюноотделения, в результате его наблюдается усиленная секреция слюны. Обильное слюноотделение происходит при ведении в кровь пилокарпина или прозерина, возбуждающих нервно-железистый аппарат слюнных желез. Наоборот, снижение или прекращение отделения слюны наступает при ведении атропина, блокирующего парасимпатическую иннервацию слюнных желез.
Определённое влияние на слюноотделение оказывают гормоны гипофиза, половых желёз, щитовидной железы, поджелудочной железы. Однако эффект их действия зависит от количества вводимого гормона.
Деятельность слюнных желез может возникать не только при раздражении рецепторов полости рта (безусловные рефлексы), но и при раздражении других рецепторных полей.
Слюнные железы, например, у взрослого человека или высших животных могут возбуждаться при виде или запахе пищи, т. е. раздражении рецепторов глаза и носа (условные рефлексы).
Возникающее при еде отделение слюны является результатом сочетания действия условных и безусловных раздражителей. Как условные, так и безусловные рефлексы строго детерминированы, т. е. всегда причинно обусловлены действием какого-либо раздражителя.
Возможно рефлекторное возбуждение слюнных желез при раздражении желудка, кишок и других полых органов (С.С.Полтырев с сотрудниками). И. П. Павлов показал, что при известных условиях раздражение нервов органов брюшной полости или седалищного нерва может повести к длительному торможению секреторной деятельности слюнных желез.
Подобный эффект наблюдается и при раздражении рецепторов кишечника.
Особенности слюноотделения и роль слюны в процессе пищеварения у жвачных животных. У жвачных животных физиология слюноотделения и роль слюны в пищеварении имеют некоторые особенности, обусловленные строением желудочно-кишечного тракта и способом питания.
При поступлении кормов в ротовую полость у жвачных животных происходит усиленное слюноотделение, способствующее смачиванию пищевых веществ. Поскольку окончательное пережевывание корма происходит не во время приема пищи, а после того, когда она вновь возвращается в ротовую полость (процесс отрыгивания) из рубца, смачивающая роль слюны приобретает особо важное значение в процессе эвакуации корма.
Слюна жвачных животных имеет высокую щелочность, что способствует нейтрализации кислот, образующихся в рубце при процессах брожения углеводов. Высокая щелочность зависит от присутствия в слюне карбонатов. Щелочность слюны у овцы равняется 0,56—0,77%, у быка — 0,62—0,65, у козы — 0,54—0,77%. Например, если учесть, что у коровы за сутки выделяется около 60 л слюны, то в рубец поступает со слюной около 300—350 г карбонатов. Плотность слюны колеблется от 1,0012 до 1,0053. Химический состав слюны у коровы, овцы, козы и некоторых других жвачных почти одинаков (табл. 5).
Таблица 5 Химический состав слюны у жвачных животных
| Вещества | Химический состав слюны, % | ||
| Корова | Овца | Коза | |
| Вода | 99,62 | 96,64 | |
| Сухие вещества | 0,38 | 0,36 | 0,71-0,96 |
| Органические вещества | 0,29 | 0,28 | - |
| Зола | 0,09 | 0,08 | 0,62-1,62 |
| Азот | 0,035 | 0,033 | 0,01-0,016 |
| Белок | 0,22 | 0,21 | - |
Диастатический фермент почти не содержится в слюне жвачных. Так, например, под влиянием слюны коровы или овцы в крахмальном клейстере в течение двух часов образуется лишь 0,38—0,75 мг сахара.
Отделение слюны у жвачных околоушными железами происходит непрерывно.
Периодическая работа в связи с приемами пищи наблюдается лишь в подъязычных и подчелюстных железах. Непрерывная секреция слюны из околоушной железы некоторыми авторами объясняется интероцептивными влияниями с преджелудков и действием некоторых гормонов. Интенсивность отделения ее зависит от процесса жвачки. В секреторном процессе околоушной железы играют определенную роль и условно-рефлекторные факторы.
Количество отделяемой слюны с переходом животного с молочного на растительный режим питания увеличивается, причем количество и качество слюны находятся в зависимости от свойств раздражителя. Сильное возбуждение секреции обусловливается пережевыванием жвачки; из концентрированных кормов овес является самым сильным возбудителем слюноотделения.
Глотание
Сложный рефлекторный акт, при котором в результате сокращения одних и расслабления других мышц пища переводится из полости рта в пищевод, а затем в желудок.
Центр глотания находится в продолговатом мозге на дне четвертого желудочка. Глотание наступает тогда, когда механически обработанная во рту пища достаточно смочена слюной.
Пищевой комок при помощи координированных движении щек и языка передвигается к глотке на корень языка, за передние дужки. При этом происходит раздражение рецепторов слизистой оболочки зева и мягкого нёба, возникшие импульсы по волокнам тройничного, языкоглоточного и верхнего гортанного нерва передаются в центр глотания. Отсюда центробежные импульсы, направляясь по двигательным веточкам тройничного, языкоглоточного, подъязычного и блуждающего нервов к мускулатуре ротоглотки, вызывают не координированные сокращения.
Глотка
Начальный отдел желудочно-кишечного тракта, соединяющий ротовую полость с пищеводом. Представляет собой воронкообразный, мышечный мешок. Стенки её состоят из трех слоев: слизистой оболочки, где находятся слизистые железы; мышечного слоя, состоящего из поперечнополосатой мускулатуры, и наружного слоя, содержащего соединительную ткань. Мускулатура глотки располагается продольно и кольцевидно.
Раздражение слизистой оболочки глотки рефлекторным путем вызывает сокращение мышц языка и мускулатуры, приподнимающей мягкое небо; благодаря этому вход в полость носа со стороны глотки закрывается мягким небом и язык перемещает пищевой комок в лотку. Одновременно происходит смещение подъязычной кости и приподнимание гортани, в результате чего надгортанник закрывает вход в гортань, предотвращая таким образом попадание пищи в дыхательные пути (рис. 11). Глотательные движения совершаются рефлекторно, как только пищевой комок достигает входа в глотку. Об этом свидетельствуют наблюдения за глотанием во время наркоза или во время сна. Смазывание слизистой оболочки глотки 3—5%-ным раствором кокаина, так же как и перерезка чувствительных нервов, например тройничного у кролика или языкоглоточного у собаки, делают глотание невозможным, что можно наблюдать и в нормальных условиях при отсутствии в ротовой полости пищи и слюны. В этом каждый легко может убедиться, если при отсутствии пищи во рту сделает несколько глотательных движений.
При нормальных физиологических условиях глотание начинается произвольно под влиянием импульсов с коры больших полушарий. В дальнейшем совершается быстрое непроизвольное передвижение пищевого комка через глотку в пищевод. Это происходит путем последовательно возникающих рефлексов («цепные рефлексы»).
Рис. 11. Схема акта глотания. А — глотка и покое, Б — глотательное движение:
1 — носовая полость; 2 — мягкое небо; 3 — язык; 4 — надгортанник; 5 — мышца дна ротовой полости; б — подъязычная кость; 7 — пищевод; 8 — гортань; 9 —пищевой комок
Деятельность центра глотания связана с деятельностью других нервных центров, расположенных в продолговатом мозге. Так, при глотании наблюдается торможение центра дыхания и возбуждение центра, регулирующего работу сердца. Поэтому при глотании происходит задержка дыхания и учащение сердечных сокращений.
Движение пищи по пищеводу. Пищевой комок из глотки попадает в пищевод, по которому не передвигается от верхнего конца к нижнему благодаря последовательным сокращениям мускулатуры пищевода, и затем поступает в полость желудка.
Рис. 12. Запись движений пищевода на лейте кимографа:
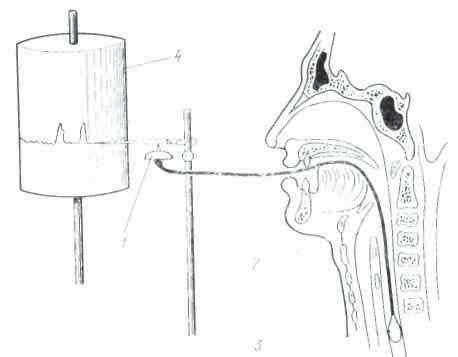
1— капсула Марея; 2 — резиновая трубка; 3 — резиновым баллон для регистрации вижений пищевода; 4 — кимограф
У человека пищевод представляет мышечную трубку длиной 25—30 см, состоящую из трех слоев: слизистого, мышечного и соединительнотканого. На всем протяжении он имеет три анатомических сужения. Первое находится на уровне задней пластинки перстневидного хряща; просвет пищевода в этом месте равен около 1,4 см. Последующие сужения находятся а уровне - бифуркации аорты и на месте прохождения пищевода через диафрагму.
Значительное сужение просвета пищевода имеется у входа в желудок из-за находящихся здесь мощных круговых мышц — кардиального сфинктера. В верхней трети мускулатура пищевода состоит из поперечнополосатых, а в остальных частях — из гладких мышц. Пищевод обладает способностью сокращаться. Наблюдения над его сокращениями производятся при помощи тонкого зонда с резиновым баллончиком на конце, который через ротовую полость вводится в пищевод. Другой конец зонда соединяется с капсулой Марея, рычажок которой записывает на кимографе сокращения (рис. 12).
Объем одновременно проглатываемой пищи приблизительно равен 5 мл. Скорость прохождения пищевого комка по пищеводу зависит от консистенции пищи. Твердая пища проходит за 8—9 с, максимум 15 с, жидкая — за 1—2 с.
Иннервация пищевода осуществляется преимущественно волокнами вагосимпатических нервных стволов: шейная часть иннервируется возвратными нервами, грудная— главными стволами блуждающих нервов, при этом правый нерв образует заднее пищеводное сплетение, левый — переднее. Оба сплетения имеют многочисленные анастомозы друг с другом и с симпатическими волокнами. Симпатические волокна происходят от шейных и звездчатых ганглиев, грудных узлов пограничного ствола и от большого чревного нерва, принимающих участие в образовании пищеводных сплетений. Пищевод получает волокна и от сплетений грудной аорты и левой диафрагмальной артерии. Что касается интрамурального нервного аппарата, то он состоит из сплетений, расположенных в адвентициальном, межмышечном и подслизистом нервных сплетениях. Пищевод—мощная рефлексогенная зона. Чувствительные нервные образования располагаются во всех его слоях и на всем протяжении. Чувствительная иннервация верхней трети пищевода (состоит из поперечнополосатой мускулатуры) осуществляется волокнами, которые проходят в стволах блуждающих нервов, на нижней трети — преимущественно волокнами, идущими к межпозвоночным ганглиям. Рецепторы его в основном относятся к механорецепторам и раздражаются проходящим по нему пищевым комком.
В момент акта глотания рефлекторно пищевод подтягивается к зеву и его начальная часть воронкообразно расширяется, принимая пищевой комок. Продвижение комка вдоль пищевода осуществляется благодаря расслаблению мышц, принимающих участие в его приеме и последующему сокращению их. Вообще же перистальтическая волна распространяется от глоточного до желудочного отверстий. Для продвижения перистальтической волны по пищеводу большое значение имеют реципрокные отношения между сокращениями продольного и кольцевого слоев мышц пищевода.
Длительные тонические сокращения мышц характерны в начальной и особенно прекардиальной частях пищевода. Быстрые сокращения и расслабления мускулатуры наблюдаются только в начальных участках пищевода, а затем периоды сокращений и расслаблений удлиняются, что, по-видимому, связано с тем, что в нижней трети пищевода преобладают гладкомышечные элементы, обладающие меньшей подвижностью, чем поперечнополосатые Скорость распространения перистальтических волн у человека 2-4 см/с.
Частота и количество перистальтических сокращений в серии могут быть неодинаковыми в разных участках пищевода (рис. 13). При наличии в пищеводе остатков пищи возникают волны сокращений, которым акт глотания не предшествовал. Это так называемые вторичные перистальтические сокращения частота которых в пищеводе собак 8—14 сокращений в 1 мин.
По данным П. Г. Богача, у голодных собак появляются периодические сокращения пищевода, возникающие и прекращающиеся одновременно с аналогичными периодами сокращений и покоя желудка.
Рис 13. Сокращения начального (Л), среднего (В) и конечного (В) отделов пищевода
Активность мускулатуры пищевода находится в определенной зависимости от состояния центральной нервной Системы. Так, например, при ее возбуждении, проявляющемся в двигательном беспокойстве, наблюдается резкое усиление сократительной активности мышц всех его отделов и, наоборот, при спокойном состоянии, дремотном и сонном, активность снижается. На сократительную активность мышц оказывает влияние ацетилхолин, в то же время адреналин заметного влияния не оказывает.
Что касается мышц кардии, то они находятся в тонизированном состоянии, препятствуя попаданию кислого содержимого желудка в пищевод. При акте глотания и прохождения по пищеводу пищевого комка тонус мышц кардии ослабляется, что позволяет комку пройти в полость желудка. Кстати, снижение тонуса кардии является активным процессом. Выявлена определенная связь тонуса мышц кардиального сфинктера с тонусом мускулатуры желудка: снижение тонуса желудка сопровождается понижением тонуса кардиального сфинктера (П. Г. Богач, С. Д. Гройсман).
Глава 5
Пищеварение в желудке
Пища, поступившая в желудок, под воздействием желудочного сока впервые подвергается значительным химическим превращениям. В зависимости от качества пища находится в желудке в течение нескольких часов; здесь она тщательно перемешивается, пропитывается желудочным соком; ее составные части, особенно белковые вещества, подвергаются расщеплению, после чего желудочное содержимое постепенно эвакуируется через привратник в двенадцатиперстную кишку.
Строение желудка
Желудок представляет собой расширенную в виде мешка часть пищеварительной трубки. В нем различают вход, или кардиа, через который, пищевая масса переходит из пищевода в желудок; тело, которое по объему равно четырем пятым всего желудка; пилороантральную часть с привратником. Последний снабжен мышечным жомом — пилорическим сфинктером, сокращение и расслабление которого обусловливают переход пищевых масс в кишечник. Кроме сфинктера в желудке, на границе между телом и привратником, имеется еще препилорический сфинктер. В теле желудка различают малую кривизну (верхняя вогнутая часть) и большую кривизну (нижняя выпуклая часть). При вертикальном положении самая высокая часть желудка, находящаяся у входа, называется входом. Емкость желудка у взрослого человека в среднем равна приблизительно 2 л, у лиц, потребляющих много жидкости, она может доходить до 5—10 л.
Стенка желудка состоит из трех оболочек: слизистой, мышечной и серозной. Слизистая, или внутренняя, оболочка выстлана однослойным, выделяющим слизь эпителием и содержит многочисленные железы трубчатой формы, которые открываются на дне желудочных ямок. В железах тела желудка (малая кривизна, дно) имеются главные, обкладочные и добавочные клетки. Главные клетки кубической формы служат для выработки фермента, в обкладочных круглой формы клетках образуется соляная кислота, добавочные клетки вырабатывают слизь. Пилорические железы содержат только главные и добавочные клетки. Следовательно, секрет, выделяемый клетками фундальной части желудка, имеет кислую реакцию, в то время как сок из пилорической части желудка не содержит соляной кислоты и обычно бывает нейтральной или слабощелочной реакции.
В желудке взрослого человека насчитывается до 25 000 000 железистых клеток.
Слизистая оболочка пустого желудка собрана в складки, которые расправляются при наполнении его пищевой массой.
Мышечная, или средняя, оболочка желудка состоит из трёх различно направленных слоев мышечных волокон: продольного, циркулярного и внутреннего косого. Благодаря такому расположению волокон желудок при сокращениях может изменять свою величину и форму во всех направлениях. Это обстоятельство обеспечивает тщательное перемешивание пищевых масс в желудке.
Серозная, или наружная, оболочка состоит из соединительнотканых волокон. Стенки желудка богато снабжены кровеносными сосудами; мелкие сосуды имеют вид спиралей или клубочков, которые при наполнении желудка растягиваются. Желудок иннервируется блуждающими и симпатическими нервами (рис. 14). Левый и правый блуждающие нервы, проходя вдоль стенок пищевода, образуют сплетение, от которого к желудку идут передний, или левый, и задний, или правый, стволы этих нервов. Оба ствола разветвляются на отдельные ветви: самая крупная ветвь идет от левого ствола вдоль малой кривизны, иннервируя при этом дно в верхние две трети тела желудка; остальные проходят по стволу тела желудка к привратнику, где одна ветвь нннервирует препилорическую часть и область привратника, а другая — направляется в печень и желчевыделительный аппарат; от последней ветви отделяются волокна, иннервирующие привратник и двенадцатиперстную кишку. От заднего ствола идут ветви по дорзальной стороне желудка; одна из них иннервирует кардию, малую кривизну и часть тела желудка; другая, небольшая, анастомозирует с правым полулунным ганглием; третья направляется к центральной части желудка.
Рис. 14. Схема иннервации желудка спереди (Л) и сзади (Б):
1 — правый вагус; 2 — левый вагус; 3 — левый чревный нерв: 4 — правый чревный нерв; 5 — аорта; 6 — ветвь левого вагуса к печени; 7 — ветвь правого вагуса к задней стенке желудка; 8 — нисходящая ветвь к привратнику; 9 — ветвь правого вагуса; 10 — ветвь правого вагуса к полулунному ганглию; 11 — ветвь правого вагуса к правому полулунному ганглию; 12 — правый чревный нерв; 13 — правый полулунный ганглий; 14 — левый полулунный англий; 15 — поджелудочная железа; 16 — нисходящая ветвь вагуса; 17 — симпатические ветви, идущие раздельно; 18 — симпатическая ветвь к кардии. Ветви вагуса окрашены серным, симпатические — серым
Оба блуждающих нерва в стенках желудка анастомозируют между собой, в результате чего каждый из них принимает участие в иннервации различных отделов желудка.
Симпатические нервы желудка идут от солнечного сплетения, в образовании которого принимают участие большие и малые чревные нервы и отчасти волокна блуждающего нерва.
Чревные нервы образуют вдоль малой и большой кривизны желудка верхнее и нижнее плетения. По последним данным, чревные нервы содержат в своем стволе парасимпатические волокна точно так же, как и блуждающие нервы имеют симпатические волокна. Таким образом, блуждающие и чревные нервы являются смешанными. В иннервации желудка принимает участие и правый диафрагмальный нерв, который имеет анастомозы с ветвями солнечного сплетения.
Рис. 15. Схема распределения нервных клеток первого и второго типов (А. С. Догель): Клетки первого типа звездчатые с отростками, второго — с одним или вумя отростками
В стенках желудка находятся два нервных сплетения: сплетение Ауэрбаха и сплетение Мейсснера.
Первое располагается между волокнами продольного мышечного слоя, второе образует несколько слоев по всей толщине слизистой оболочки; оба сплетения соединяются между собой (рис. 15). По данным некоторых авторов, у человека и собаки имеется еще третье сплетение— на наружной поверхности желудка, непосредственно под брюшиной.
В области кардии, привратника и малой кривизны нервная сеть плотнее, а отдельные ганглии крупнее, чем в районе дна и большой кривизны желудка. В среднем на см2 поверхности стенки желудка приходится в области дна — от 80 до 200, в области тела — от 250 до 320 и в области привратника— до 450 нервных клеток.
Мейсснеровское сплетение состоит из двух отдельных образований: поджелезистого, или подслизистого, и межжелезистого, нервные волокна которого окружают желудочные железы. Оба эти образования соединяются между собой при помощи длинных, узловатой формы волокон, причем многие нервные волокна подслизистого сплетения начинаются от леток ауэрбаховского сплетения, особенно в области привратника и малой кривизны.
Согласно гистоморфологическим исследованиям Б. И. Лаврентьева, Н. Г. Колосова и р., в желудке имеются нервные клетки первого типа, которые тесно связаны с ветвями блуждающих нервов и имеют отношение к двигательной функции желудка.
В желудке находится рецепторный аппарат, сигнализирующий в высшие отделы центральной нервной системы 0 степени растяжения желудка — механорецепторы или барорецепторы, о функциональном состоянии; его мускулатуры — проприорецепторы, о химических изменениях содержимого желудка — хеморецепторы, об изменении температуры — терморецепторы и об изменении осмотического давления в крови и тканях желудка — осморецепторы. Афферентными проводниками импульсов, возникающих в рецепторах желудка, являются волокна как блуждающих, так и чревных нервов. Таким образом, блуждающие и черепные нервы желудка являются одновременно и афферентными нервами.
Основные секреторные нервы желудка — блуждающие нервы, при раздражении которых возникает секреция желудочного сока, а кроме того, они возбуждают моторную деятельность желудка. В составе блуждающих нервов имеются и тормозящие волокна, раздражение которых угнетает деятельность секреторных клеток. При раздражении чревных нервов происходит торможение моторики желудка. После перерезки блуждающих нервов раздражение чревных нервов вызывает секрецию желудочного сока (Г. В. Фольборт).
Методы изучения желудочной секреции
Изучение желудочной секреции у животных производится в хроническом опыте после проведения специальных операций, позволяющих получить секрет желудочных желез. У собак и некоторых сельскохозяйственных животных (лошадь, корова, свинья, овца и др.) проделываются следующие операции: наложение хронической фистулы желудка, перерезка пищевода (эзофаготомия) в сочетании с наложением фистулы желудка, образование изолированного желудка по Павлову, Клсменциеви-чу-Гейденгайну, Савичу — Бресткину, изоляция пилороантралыюй части желудка с наложением гастроэнтероанастомоза, изоляция привратника без сохранения и с сохранением иннервации и др.
Хроническая фистула желудка. С экспериментальной целью фистула желудка собаке была впервые наложена в 1842 г. В. А. Басовым. С некоторыми видоизменениями эта операция производится и в настоящее время. Она заключается в том, что собаке под общим наркозом, с соблюдением всех правил асептики и антисептики, в переднюю стенку желудка вставляется и фиксируется специальная трубка из нержавеющего металла, один конец которой с фиксированной шайбой открывается в полость желудка, а другой выводится наружу. После выздоровления животного достаточно открыть пробку трубки, чтобы получить желудочное содержимое, где обычно кроме желудочного сока находятся остатки пищи и слюна. Для получения чистого желудочного сока к фистуле желудка добавляют перерезку пищевода (эзофаготомия).
Одновременное наложение фистулы желудка и перерезка пищевода было впервые произведено И. П. Павловым совместно с Г. О. Шумовой-Симановской в 1889 г.
Эзофаготомия производится в шейной части пищевода, причем последний перерезывается полностью или частично, с оставлением участка па задней стенке. Перерезанные или надрезанные концы пищевода пришиваются к кожной ране. У животного после такой операции во время еды проглоченная нища вываливается наружу через верхнее отверстие перерезанного пищевода, т. с. происходит мнимая еда. Питание собак после опыта производится путем вкладывания пищи в желудок через желудочную фистулу или через нижнее отверстие перерезанного пищевода. Гастроэзофаготомированные собаки в соответствующих условиях могут жить годами, мало чем отличаясь по состоянию здоровья от неоперировапных животных.
Во время мнимой еды из фистулы желудка выделяется чистый желудочный сок, который можно измерить и проанализировать. Этот опыт И. П. Павлов назвал опытом «мнимого кормления» (рис. 16). Таким образом, собаки могут часами есть пищу, совершенно не насыщаясь ею, а за это время у них выделяется до 1 л чистого желудочного сока высокой кислотности и переваривающей силы.
Этот опыт натолкнул И. П. Павлова на мысль использовать натуральный желудочный сок собаки в клинической практике для лечения больных, страдающих недостаточной функцией желудочных желез (анацидный и гипоацидный гастрит и др.). В 1890 г. И. П. Павловым была организована «фабрика желудочного сока», которая до сих пор снабжает медицинские учреждения высококачественным лечебным препаратом. Применяемый в этих случаях желудочный сок собак проходит предварительную обработку и фильтрацию через специальные мелкопористые фильтры, задерживающие микробы. Разлитый в стерильную посуду сок сохраняет целебные свойства в течение многих месяцев. Клиническая практика свидетельствует о его лечебных свойствах не только при заболеваниях желудка и других органов пищеварения, по и при заболеваниях кроветворных органов (анемия).
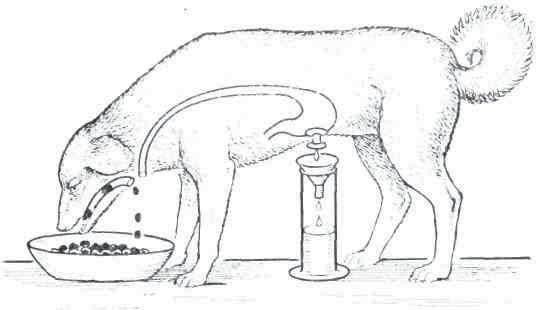
Рис. 16. Схема опыта мнимого кормления собаки с фистулой желудка и эзофаготомией
Гастроэзофаготомия по Павлову позволила изучить влияние акта еды на работу желудочных желез, проанализировать начало и развитие секреторного процесса и установить роль блуждающих нервов как секреторных нервов желудка.
Для изучения секреторного процесса в желудочных железах во время пищеварения И. П. Павлов (1894) разработал оригинальную операцию образования изолированного маленького желудочка, разобщенного с полостью большого желудка и имеющего с ним связь только через сосуды и нервы.
Операция производится следующим образом. Из стенки фундаль-ной части желудка выкраивается лоскут, причем серозная и мышечная оболочки в месте соединения маленького желудочка с большим желудком сохраняются, слизистая перерезается и из нее делается свод, отделяющий полость большого желудка от изолированного маленького желудочка. Края последнего сшиваются, наружное отверстие его пришивается к краям кожной рапы. На перерезанные стенки большого желудка накладываются швы (рис. 17).
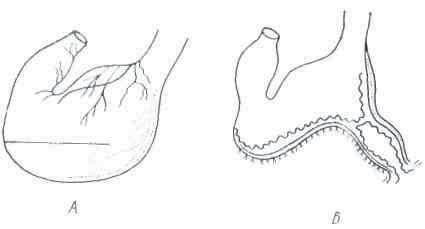
Рис. 17. Схема операции образования изолированного желудочка (по И. П. Павлову). А — линия разреза, Б – изолированный желудочек
Таким образом, при нахождении пищи в большом желудке из изолированного маленького желудочка можно получить чистый желудочный сок, по количеству и качеству которого можно определять течение секреторного процесса в желудочных железах. Благодаря сохранению иннервации в маленьком желудке работа желез последнего, как в зеркале, отражает работу секреторных клеток всего желудка (табл. 6).
Изолированный маленький желудочек по Павлову принципиально отличается от изолированного маленького желудочка по Клеменциевичу—Гейденгайну тем, что при выкраивании последнего перерезываются нервы и, следовательно, исключаются нервные влияния на дея тельность желудочных желез изолированного желудочка.
Тавлица 6. Секреция желудочного сока из большого желудка и маленького желудочка у гастроэзофаготомированнои собаки при мнимом кормлении (по И. П. Павлову)
| Часы | Маленький желудочек | Большой желудок | ||
| количество сока, мл | переваривающая сила, мм | количество сока, мл | переваривающая сила, мм | |
| 1 2 3 | 7,6 4,7 1,1 | 5,88 5,75 5,5 | 68,25 41 ,5 14,0 | 5,5 5,5 5,38 |
| Средняя величина | 13,4 | 5,75 | 123,75 | 5,5 |
Г.М. Давыдовым была разработана операция наложения изолированного маленького желудочка на малую кривизну желудка. Технически она проводится так же, как и операция наложения маленького желудочка на большую кривизну. В последнее время Л. В. Соловьев для изучения секреторной реакции желудка у собак предложил накладывать одновременно на малой и большой кривизне изолированные желудочки по Павлову.
Значение этой методики состоит в том, что она позволяет изучить особенности секреторного процесса различных секреторных полей желудка, которые, как показали эксперименты, неоднозначны. Кроме того, изучение функции желез малой кривизны желудка имеет большое клиническое значение, так как именно в этом месте значительно чаще, чем в других областях желудка, возникают язвенный процесс и злокачественные новообразования.
Павловский метод хронических фистул был широко использован для изучения физиологии желудка у сельскохозяйственных животных. Уже в 1898 г. И. В. Рязан-цев впервые оперативным путем выкроил у быка изо-лированный желудочек, а в 1910 г. В. В. Савич и Н. П. Тихомиров провели исследование па козе с изолированным по Павлову желудочком. То же произвел и И. В. Бельговский в 1912 г. на телятах, а Н. Ф. Попов в 1937 г. на лошадях.
Работами советских ученых были детально изучены секреторные и двигательные процессы у птиц и у различных видов животных с хроническими фистулами желудка.
Изучение желудочной секреции у человека имеет важное диагностическое значение при заболеваниях желудка (гастрите, язве, раке), печени, поджелудочной железы, кишечника, а также органов мочеотделительной, сердечно-сосудистой и других систем организма. В клинике принято проводить исследование желудочной секреции при помощи толстого или тонкого резинового зонда, вводимого через рот в желудок.
Для возбуждения желудочных желез применяют различные пищевые или химические вещества, так называемые пробные завтраки: уха — по Горшкову, капустный сок — по Лепорскому, пиво — по Михайлову, раствор алкоголя по Эрмапу, сухари с чаем — по Боас-Эвальду и т. д.
Определение желудочной секреции при помощи методики толстого зонда не дает точных данных, так как из желудка получается не чистый желудочный сок, а желудочное содержимое, состоящее из неизвестного количества желудочного сока и введенного пробного завтрака. К тому же желудочное содержимое берется одномоментно, только в разгар пищеварения. Методика тонкого зонда более ценна, так как позволяет исследовать деятельность желудочных желез в течение 2—3 ч, что дает представление о динамике секреторного процесса. Недостаток ее заключается также в том, что в большинстве случаев получаемое содержимое желудка— смесь желудочного сока с введенным пробным завтраком. Для получения чистого желудочного сока в клинике Н. И. Лепорского была разработана методика получения желудочного сока у человека при приеме 10 мл 33%-ного раствора алкоголя. Получаемый в этих случаях при помощи тонкого зонда желудочный сок свободен от пищевых веществ и поэтому анализ его проводится по абсолютным цифрам кислотности и пе-реваивающей силы Кроме того, в соке можно точно определять количество лейкоцитов, поступающих в желудочный сок из слизистой оболочки желудка (лейкопедез). Определение лейкопедеза в желудочном соке больных имеет важное диагностическое значение, особенно при раке, гастрите и язве желудка.
Значительное отделение желудочного сока наблюдается при внутримышечном или подкожном введении солянокислого или фосфорнокислого гистамина. Иногда введение гистамина комбинируют с фракционным методом исследования, вводя в желудок пробный завтрак. Обыкновенно это практикуется в том случае, когда в полученных первых порциях желудочного содержимого отсутствует свободная соляная кислота. Метод гиста-миновой стимуляции желудочных желез применяется главным образом у больных, когда требуется выяснить, является ли отсутствие свободной соляной кислоты в желудочном соке результатом чисто функционального нарушения деятельности секреторных клеток или оно обусловлено органическим (структурным) изменением последних.
В некоторых клиниках применяют инсулиновый метод возбуждения желудочных желез. С этой целью больному вводят подкожно или внутривенно 12—20 единиц инсулина; отделяющийся после этого желудочный сок извлекают при помощи тонкого зонда.
Все эти методы, основанные на гуморальной стимуляции желудочных желез, точно так же, как и введение пробного завтрака через зонд в желудок, позволяют определить его только в период нервно-химической фазы секреции, что значительно снижает их теоретическую и практическую ценность, так как для диагностики представляется важным определение функций желудочных желез и в первой сложнорефлекторной фазе секреции.
В последнее время был разработан новый метод получения чистого желудочного сока у человека при помощи раздражения механорецепторов желудка (И.Т.Курцин и Н. Е. Слунский). Для этого был сконструирован желудочный зонд, состоящий из двух тонких резиновых трубок, па конце одной из этих трубок имеется тонкий каучуковый баллончик, который вводится в желудок и раздувается 250 мл воздуха; в таком состоянии баллончик находится в желудке в течение 1—2 ч. Образующийся при раздражении механорецепторов желудка сок откачивается через вторую трубку зонда, а затем подвергается точному количественному и качественному анализу. Этот метод был усовершенствован таким образом, что кроме определения работы желудочных желез в сложнорефлекторную фазу производится определение секреторной функции желудка и во вторую нервно-химическую фазу секреции (К. М. Быков и И.Т.Курцин). После часового раздражения механорецепторов желудка и получения сока в период первой фазы секреции воздух из баллончика удаляется, а в желудок через вторую трубку зонда вводится жидкий пробный завтрак (капустный сок, раствор алкоголя и т. п.), затем в течение часа через 15-минутные интервалы извлекаются отдельные порции желудочного содержимого для анализа. Дальнейшая модификация этого метода позволила исследовать у человека не только секреторную, но и двигательную функцию одновременно, для чего трубка зонда, снабженная баллончиком, соединяется через водяной манометр с капсулой Марея, рычажок которой непрерывно записывает на кимографе сокращения стенок желудка (рис. 18). Кроме того, определение функции желудочных клеток ко вторую фану секреции, так же как и в черную, производится на ос-новании получения чистого желудочного сока. Это достигается тем, что через 30 мин после введения пробного завтрака последний удаляется и в течение последующего часа из желудка извлекается через каждые 15 мин последовательный сок, который почти не содержит примеси пробного завтрака и таким образом является чистым секретом желез.

Рис. 18. Метод комплексного изучения секретоной и двигательой функции желудка у человека (по П. Г. Курнношу)
Кроме описанных методов зондирования, широко применяемых в клинической практике, изучение желудочной секреции у человека удавалось еще и в тех случаях, когда вследствие травматического повреждения ж.елудка после ранения или оперативного вмешательства, вызванного непроходимостью пищевода, образуются свищи желудка. Встречаются иногда и больные с га-строэзофаготомией. В литературе описан случаи, когда у человека после воспалительного процесса образовался в дальнейшем изолированный маленький желудочек. Физиологические наблюдения, проведенные на таких больных, представляют исключительный теоретический и практический интерес, так как полученные данные составляют основу подлинной физиологии желудка человека. Встречаются иногда и больные с га-строэзофаготомией. В литературе описан случаи, когда у человека после воспалительного процесса образовался в дальнейшем изолированный аленький желудочек. Физиологические наблюдения, проведенные на таких больных, редставляют исключительный теоретический и практический интерес, так как полученные анные составляют основу подлинной физиологии желудка человека.
Состав и пищеварительные свойства желудочного сока
Суточное количество сока у человека 1,5—2,5 л, у собаки — 2,3 л, у лошади и коровы — 10—30 л. При смешанной еде у человека за один прием пищи отделяется от 700 до 800 мл сока.
Чистый желудочный сок человека и животных (собаки, кошки) представляет собой бесцветную, слегка опалесцирующую жидкость кислой реакции. В его состав входят вода, соли, ферменты, слизь и кислоты. Кислая реакция сока зависит главным образом от присутствия свободной соляной кислоты, содержание которой равно у человека 0,4—0,5% и у собаки 0,5—0,6°/о. По Павлову, снижение кислотности желудочного сока зависит от нейтрализующего действия щелочного сока пилорических желез и щелочной слизи, отделяемой слизистыми железами. Поэтому у человека общая кислотность желудочного сока, получаемого при помощи зонда, обычно бывает равной 0,14 0,25%, а свободная соляная кислота — 0,07—0,13%; рН чистого желудочного сока человека составляет 1,0 — 1,5. Плотный остаток сока колеблется около 0,3—0,4%- Плотность равна 1,0083—1,0086. Из основных неорганических веществ в состав желудочного сока человека входят хлориды, фосфаты, азотнокислые соли, железо; из органических-молочная, пировиноградная и креатинфосфорная кислоты, сахар, иуклеопротеиды, ферменты. При микроскопическом анализе в соке обнаруживаются крахмальные зерна, мышечные волокна, растительные клетки, эпителий, эритроциты, лейкоциты, микробы, дрожжевые грибки, сарцины и др.
Согласно современным данным, в желудочном соке содержатся четыре желудочных фермента. Группы пептид-пептидогидролаз: пепсин, гастриксин, пепсин В и реннии. Пепсин переваривает белки, однако не оказывает гидролитического действия на муцин и кератин волос. Гастриксин является ферментом желудочного сока человека и обладает максимально высокой протео-лптической активностью (при рН 3,2). Он активнее пепсина и способен расщеплять гемоглобин, однако уступает ему в скорости гидролиза яичного белка. Пепсин В и парапепсин имеют сходство в транспептидазном действии, причем оба угнетаются при рН 5,6. Репнин (сычужный фермент, или химозин) является характерным компонентом желудочного сока молодых жвачных животных. По специфичности он близок к пепсину. Этот фермент относится к коагулирующим ферментам из-за способности свертывать молоко. Кроме того, он способен инактивировать рибонуклеазу.
В составе желудочного сока содержатся и непротео-литическне ферменты, в частности лизоцим, который вырабатывается, по-видимому, клетками поверхпостно-го эпителия и придает ему бактерицидные свойства. Желудочный сок обладает липо- и амилолитической активностью. Предполагают, что липаза и амилаза сек-ретируются из крови пилорическими железами (Г. Ф. Коротько с сотрудниками).
Физиологическое значение соляной кислоты желудочного сока заключается в следующем: 1) она активатор пепсина, который переваривает белки только в кислой среде; 2) под ее влиянием разбухают белки пищи, что содействует их перевариванию; 3) действуя па слизистую пилороантральной части желудка, переводит из неактивного состояния в активное гормон гастрин, участвующий в возбуждении фундальных желез желудка; 4) поступая в двенадцатиперстную кишку, стимулирует образование гормона секретина, возбуждающею поджелудочную секрецию; 5) рефлекторно вызывает со слизистой двенадцатиперстной кишки сокращение пило-рического сфинктера; 6) оказывает бактериостатическое и бактерицидное действие на микробы, поступающие с пищей в желудок; 7) декальцинирует и тем самым размягчает кости.
Диализ кислотности желудочного сока производится при помощи титрацнопного способа или электрометрическим способом (с помощью рН-метра).
Физиологическое значение слизи состоит в том, что она: 1) играет защитную роль, предохраняя слизистую желудка от вредного действия механических и химических раздражителей (И. П. Павлов); 2) активный пищеварительный агент, так как абсорбирует ферменты желудочного сока (М. П. Бресткин, К. М. Быков, Л. В. Риккль); 3) присоединяет растворимые в воде витамины В и С, благодаря чему они не разрушаются желудочным соком (И. П. Разенков); 4) содержит свободные вещества, обладающие способностью возбуждать секреторную функцию желудка (И. П. Разенков).
Желудочный сок малой кривизны желудка. По данным лаборатории К- М. Быкова, желудочный сок, выделяемый клетками малой кривизны, обладает повышенными переваривающими свойствами и кислотностью. Отделение его во время приема пищи начинается значительно раньше по сравнению с другими секреторными полями. Секреция сока обычно в первые часы пищеварения очень интенсивная и прекращается при наличии еще продолжающейся секреции на большой кривизне желудка. По К. М. Быкову, малая кривизна является ведущей частью в развитии секреторного процесса всего желудка. Структурные изменения, наблюдаемые в секреторных клетках при длительной секреции, возникают главным образом в зоне малой кривизны желудка (И. П. Разенков).
В отличие от фундальных желез и желез малой кривизны, выделяющих кислый сок, пилорическая часть желудка выделяет привратниковый сок щелочной реакции. Как показали опыты лаборатории И. П. Павлова, отделение привратникового сока происходит непрерывно, даже вне пищеварения. Механические и некоторые химические раздражители усиливают его секрецию. Сок содержит пепсин, слабо действующий в щелочной среде, в большом количестве слизь, воду, соли, главным образом углекислые, и некоторые белковые вещества. В привратниковом соке находится особый глико-литнческий фермент, расщепляющий гликоген до молочной кислоты (И. П. Разенков). С началом секреторного процесса и особенно в разгар его желудочный сок, благодаря появлению в нем холипергпческого медиатора, приобретает холипергические свойства (Я- П. Скляров).
Бактерицидные и гемопоэтические свойства желудочного сока. Желудочный сок обладает высокими бакте-риостатическими и бактерицидными свойствами благодаря наличию в нем свободной соляной кислоты и особого ферментоподобного вещества лизоцима. Так, например, гноеродные бактерии (стафилококки, стрептококки и др.) погибают в нем через 2—4 мин, холерные вибрионы — через 10—15 мин.
Желудочный сок человека содержит вещество гема-мии, пли фактор Кастля, который способствует усвоению витамина В12, необходимого для нормального созревания эритробластов. Выяснено, что химическая структура этого противоанемического вещества соответствует структуре витамина B12, в состав которого входит кобальт. К этому следует добавить, что слизистая оболочка желудка, так же как костный мозг, кишечник, селезенка и печень, является депо ферритина — белкового соединения железа, участвующего в синтезе гемоглобина. Вот почему у некоторых больных после резекции желудка часто развивается анемия. Образующееся в стенках желудка противоанемическое вещество депонируется печенью. Поэтому в клиниках больных, страдающих анемией, лечат или кормлением сырой печенью, или введением печеночных препаратов (компалона). Противоанемпческое вещество содержится и в желудке свиньи, у собак, оно, по данным некоторых авторов, выделяется только пплороантралыюй частью.
По данным лаборатории II.П.Разепкона, у собак через 12—16 месяцев после удаления пилорической части желудка фундальные железы начинают продуцировать сок, стимулирующий кроветворение (гемопоэз).
Экскреторная функция желудка
В желудочном соке обнаружен ряд веществ, которые выделяются из организма в виде экскретов (мочевина, мочевая кислота, креатинин и др.). Выделение этих метаболитов с желудочным соком значительно повышается при заболеваниях почек (Р.Л. Лурия и др.), органов дыхания (С.С. Полтырев, А.А. Шарыгин, Н.М. Сковородни и др.; А.Я. Губергриц, Б.Д. Боревская и др.), органов пищеварения (С.С. Полтырев с сотрудниками и др.). Некоторые химические вещества, введенные парентерально, выделяются слизистой желудка. К таким веществам относится ряд красок (нейтральрот, метплен-блау и др.). На этом свойстве слизистой желудка основан метод хромоскоппи желудка, имеющий диагностическое значение (Р.А. Лурпя).
Человеку вводят под кожу или внутримышечно 2 мл 1%-ного водного раствора нейтральрота и одновременно через заранее введенный в желудок зонд вводится какой-либо пробный завтрак (спиртовой, кофеиновый и т. п.). Извлекая затем через каждые 2—3 мин желудочное содержимое, отмечают момент появления жидкости красноватой окраски. У здорового человека эта окраска, появляющаяся в связи с выделением нейтральрота, обнаруживается через 12— 18 мин.
По мнению многих авторов, выделение нейтральрота происходит обкладочными клетками фундальнон части желудка. Метод хромоскоппи модифицирован и усовершенствован А. Е. Гельфманом. Недавно было показано, что в экскреции нейтральрота принимает участие не только желудок, но и печень. После инъекции краска обнаруживалась в желудочном соке на 5-й мин., в желчи— на 13-й мин. Максимальная концентрация нейтральрота в желудочном соке падала на первый час секреторного периода, а в желчи — на второй. Эти закономерности сохраняются независимо от способа воз-, буждения желудочных желез и печени (мнимое кормление, введение гистамина). Эти новые данные свидетельствуют о том, что экскреторные процессы в желудке и печени взаимосвязаны (С.С.Полтырев, Р. Я- Сафи-ханов). По данным И. П. Разенкова, желудок, кишечник и печень при длительном голодании способны выделять с желудочным соком значительные количества белка (альбумины, глобулины крови), который под влиянием пищеварительных соков превращается в аминокислоты, последние, всасываясь, идут па пополнение запасов клеток и тканей. Биологический смысл этого явления состоит в том, что при длительном голодании организма альбумины, глобулины, полипептиды, находясь в крови, не могут быть использованы клетками, которые способны лишь утилизировать конечные продукты распада белка — аминокислоты. Выделяясь с пищеварительными соками в полость желудка и кишечника, белок подвергается расщеплению ферментами до аминокислот, которые затем всасываются в кровь и используются жизненно важными органами и тканями для энергетических и синтетических процессов.
Возбудители желудочной секреции. Вне пищеварительного периода у человека и некоторых животных (собака, кошка) железы дна и тела желудка не выделяют кислого желудочного сока, отделяется лишь слизь нейтральной или слабощелочной реакции.
Во время акта еды и при поступлении пищи в желудочно-кишечный канал клетки дна и тела желудка отделяют сок. Начало отделения сока не совпадает с моментом начала еды, должно пройти от 5 до 10 мин (латентный период), прежде чем появятся первые капли секрета. При еде белковой пищи (мясо) латентный период сокоотделения в среднем равен 8 мин, при углеводной (хлеб)—63/4 мин и при жирной (молоко) — 9 мин.
Латентный период сокоотделения в различных отделах желудка бывает неодинаков. Железы малой кри визны начинают выделять сок значительно раньше, чем железы дна и тела желудка (табл. 7).
Таблица 7. Величина латентного периода желудочной секреции на большой и малой кривизнах желудка при приеме различныхсортов пищи (по Г. М. Давыдову)
| Сорт пищи | Величина латентного периода | |
| большая кривизна | малая кривизна | |
| Молоко (600 г) Мясо (250 г) Хлеб (250 г) | 8 мин 5 мин 7 мин | 3 мин 15 с 3 мин 30 с 3 мин 20 с |
Количество выделяемого во время пищеварения сока прямо пропорционально количеству принятой пищи, но при большом увеличении количества пищи эта пропорция не сохраняется. Ход и продолжительность секреции, а также качество выделяющегося сока неодинаковы при приеме разных сортов пищи, поэтому II. II. Павлом стал применят!) для изучения работы желудочных желез рапные по содержанию азога основные три сорта пищи: белковую (100 г мяса), жировую (000 г молока) и углеводную (250 г хлеба). В среднем продолжительность секреторного периода на 100 г мяса равняется 8 ч, па 600 г молока — 6 ч и на 250 г хлеба 10 ч (табл. 8).
Таблица 8. Секреция желудочного соки при приеме различных сортов пищи (по И.П.Павлову)

При приеме мяса максимальное отделение сока падает на первые два часа: соответственно этому и ход кривой секреции характеризуется резким подъемом в начальном периоде сокоотделения, достигая максимума в первом или по втором часу, а затем кривая постепенно падает и довольно скоро сходит на пет. При приеме хлеба максимальное отделение падает па первый час. Кривая секреции характеризуется резким подъемом в первый час с последующим крутым падением и длительным течением па низком уровне. При приеме молока кривая секреции постепенно поднимается вверх, достигая максимума в большинство случаен на третьем часу (рис. 19).
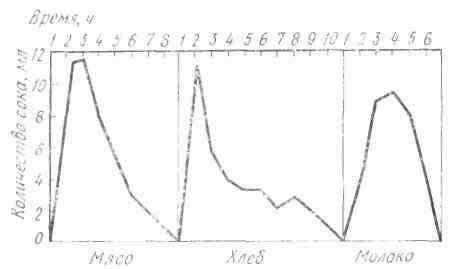
Рис. 19. Ход отделения желудочного сока у собаки при кормлении мясом, хлебом, молоком (по II. П. Павлову)
Фазы желудочной секреции
Как показали исследования лаборатории И. П. Павлова, секреторный процесс в желудке протекает в две фазы: сложнорефлекторную и нервно-химическую.
Сложнорсфлекторная фаза обусловлена возбуждением желез во время акта еды, причем возбуждение осуществляется как по типу врожденной, безусловно-рефлекторной реакции, так и по типу условнорефлекторной.
Опытами па животных было установлено, что возбуждение желудочных желез в период первой, сложно-рефлекторной фазы секреции является рефлекторным актом, причем отделение сока при виде и запахе пищи, а также звуках, связанных с приемом пищи, осуществляется путем условного рефлекса, а при акте еды — путем условного и безусловного рефлексов. Воспринимающими поверхностями для условного рефлекса являются рецепторы глаза, носа и уха, а для безусловного — рецепторы слизистой рта и глотки. Афферентными проводниками импульсов служат многочисленные нервы: вкусовые, зрительные, обонятельные, слуховые и др. Рефлекторная дуга проходит через центры продолговатого мозга, где берут начало блуждающие нервы.
При условнорефлекторном возбуждении желудочных желез человека и высших животных дуга рефлекса обязательно проходит через кору больших полушарий. На человеке это было установлено при исследовании больных с фистулами людях с неповрежденным желудочно-кишечным трактом (рис. 20).
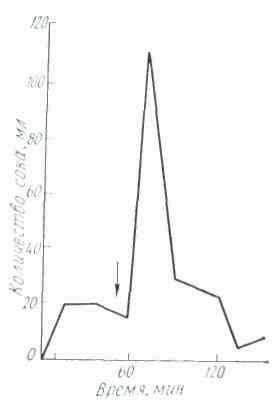
Рис 20. Условнорефлекторное отделение желудочного сока желудка и с изолированным у человека (по И.Т.Курцину желудочком, а также на и П. Е. Слуискому): Стрелкой указан момент раздражения
Желудочный сок, отделяющийся на вид и запах пищи, обычно по качественному составу соответствует тому пищевому раздражителю, которым он был вызван. Так, при виде молока отделяется сок, бедный ферментами; при виде хлеба — богатый ими. При гипнотическом внушении человеку, что он ест мясную котлету, происходит отделение сока, подобного количественно и качественно соку, отделяющемуся при еде мяса (рис. 21).
Желудочный сок, выделяемый при виде и запахе пищи, И. П. Павлов назвал «запальным» или «аппетитным» соком. Его физиологическое значение состоит в том, что желудок заранее подготавливается к перевариванию пищи.
Сильное возбуждение желудочных желез происходит при акте еды. Впервые это было установлено И. П. Павловым и Е. О. Шумовой-Симановской в 1889 г. в опыте «мнимого кормления». Ими было показано, что после начала мнимого кормления выделение первых капель сока происходит в среднем через 5 мин, продолжительность сокоотделения при 5-минутном мнимом кормлении равняется 2—4 ч; количество выделившегося сока всегда при этом бывает значительно большим, чем при раздражении животного видом и запахом пищи. Подобные результаты были получены рядом исследователей и па людях, которым по медицинским показаниям были сделаны эзофаготомия и фистула желудка; мнимая еда вызывала у них многочасовое отделение чистого желудочного сока с высокой кислотностью и переваривающей силой.
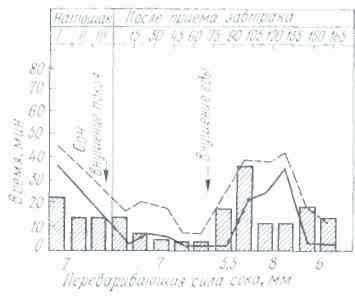
Рис. 21. Секреция желудочного сока у человека при гипнотическом внушении, что он находится в покое или ест бифштекс (по И. II. Разенкову): Сплошная кривая — свободная соляная кислота, прерывистая — общая кислотность
У собаки при полном выключении акта еды, что достигается непосредственным введением пищи через фистулу в желудок, секреция желудочного сока начинается только через 30 мин, а затем она медленно возрастает, достигая максимума лишь во втором часу; общее количество выделенного сока, так же как и переваривающая сила, в 2—3 раза меньше, чем при нормальном приеме такого же количества пищи; несколько снижается и кислотность.
И. П. Павлов сложил по часам количества сока, выделившегося при мнимом кормлении животного мясом и при введении мяса непосредственно в желудок, и построил суммарную кривую секреции желудочного сока.
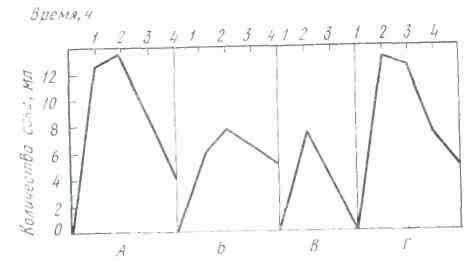
Рис. 22. Кривые секреции желудочного сока у собаки (по И. П. Павлову). А — при еде 200 г мяса, Б — при вкладывании 150 г мяса в желудок, В — при мнимом кормлении мясом, Г — кривая, получающаяся при сложении Б и В
Оказалось, что тип суммарной кривой весьма схож с типом кривой секреции, полученной при еде мяса (рис. 22). Точно так же близкими оказались и валовые количества сока, полученные таким образом.
Находясь в желудке, пища, благодаря присущим ей физическим свойствам (объем, консистенция, температура), раздражает механо- и терморецепторы, заложенные в стенках желудка, в результате чего возникает рефлекторное отделение желудочного сока. Для возбуждения секреции желудочного сока па механическое раздражение необходимы следующие условия: 1) определенный оптимум возбудимости желудочных желез, 2) длительное раздражение возможно большей поверхности слизистой желудка и 3) определенная сила раздражения.
У человека латентный период сокоотделения при раздражении механорецепторов равен 5 мин. Характер кривой секреции зависит от .вида механического раздражителя. При раздражении стенок желудка резиновым баллоном, имитирующим растяжение желудка поступившей в него пищей, ход кривой секреции сока вначале напоминает кривую секреции на мясо и хлеб. У здорового человека на этот вид раздражения отделяется в среднем около 100 мл сока за 1 ч. При повышенной возбудимости нервно-железистого аппарата желудка у человека можно получить при раздражении механо-рецепторов желудка в течение 3 ч до 1 л чистого желудочного сока, имеющего наивысшую кислотность, равную 0,49%, и переваривающую силу, равную в среднем 15 мм. Сок бывает богат слизью особенно при грубом механическом раздражении. Одновременное раздражение механорецепторов и терморецепторов желудка резко усиливает секрецию желудочного сока. Исследованиями, проведенными на больных с фистулой и эзофаготомией, было установлено, что раздражение механорецепторов желудка путем растяжения его стенок баллоном резко повышает активность желудочных желез, вызванную мнимой едой. По биохимическим свойствам желудочный сок, полученный при раздражении механорецепторов желудка, почти идентичен соку, полученному при раздражении желудочных желез пищевыми веществами (табл. 9).
Предварительное раздражение механорецепторов желудка повышает возбудимость желудочных желез к последующему действию химических и пищевых раздражителей.
Механический фактор играет существенную роль в секреторной функции желудочных желез не только у человека, но и животных. По данным С. И. Чечулина, механическое раздражение желудка вызывает у собак отделение сока в среднем через 30—45 мин после начала раздражения. Отделяющийся сок идентичен желудочному соку, выделяющемуся при приеме пищи.
Таблица 9. Количество и качество чистого желудочного сока у человека при мнимом кормлении и раздражении мсханорецепторов стенок желудки (по И.Т.Курцину и II. Е. Слупскому)
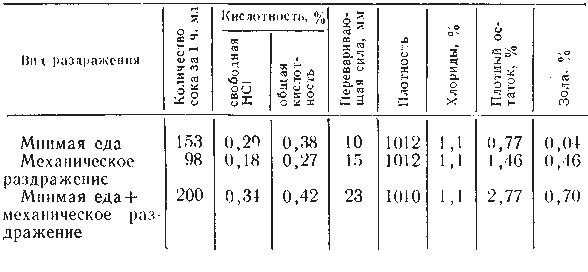
Механическое раздражение желудка вызывает секрецию желудочного сока также у черепахи, лягушки, кошки, свиньи, лошади и птиц. У некоторых животных роль механического раздражителя в процессе возбуждения желудочных желез настолько велика, что дает основание считать этот вид раздражения основным возбудителем секреции.
Пищевые и химические вещества развивают свое сокогонное действие при поступлении в привратниковую область желудка и верхние отделы тонкого кишечника. Опытами было установлено, что ряд химических агентов (вода, поваренная соль, сода, соляная кислота), экстрактивные вещества мяса и рыбы, продукты переваривания растительных и животных белков, а также некоторые секреты пищеварительных желез (слюна, поджелудочный сок, желчь), введенные на 2-3 ч через фистулу в обособленную фундальную часть желудка, не вызывают секреции желудочного сока. Введение этих же веществ в привратниковую область желудка вызывает энергичное отделение желудочного сока из фундальной части. Подобный эффект наблюдается и при раздражении мехапорецепторов привратника. Другие вещества, как, например, крепкие растворы поваренной сели, 0,5%-ный раствор глюкозы, жир, сода, олеиновая кислота, при введении в двенадцатиперстную кишку оказывают тормозящее действие на секреторную функцию желудочных желез. Особый интерес в этом отношении представляет жир и продукты его переваривания. В работе желудочных желез при жирной пище различают две противоположные фазы. В первой фазе, совпадающей с поступлением жира в двенадцатиперстную кишку, жир угнетает секреторный процесс желудочных желез. Эта фаза, в зависимости от количества введенного жира, продолжается от 2 до 4 ч. Во второй фазе, продолжающейся также несколько часов, начинается более пли менее значительное отделение желудочного сока, обусловленное действием продуктов расщепления жира (мыла, олеиновая кислота) на слизистую привратника. Естественно, что жир, обладая таким специфическим действием, при добавлении к другим сортам пищи в значительной степени изменяет ход секреторной реакции на эти вещества: латентный период возрастает, количество отделяемого сока падает, продолжительность секреторного периода и переваривающая сила сока снижаются. При этом особенно сильно тормозится деятельность желудочных желез малой кривизны (табл. 10). Поэтому при составлении меню обычно к жирной пище добавляются такие пищевые вещества, которые вызывают интенсивную секрецию желудочного сока, например различные овощи и их соки (капустный, свекольный и т. п.).
Таблица 10. Тормозящее влияние жира на секрецию из изолированных по Павлову желудочков на большой и малой кривизнах желудка
| Сорт пищи, г | Объем желудочной секреции, мм за 5 ч | |
| алая кривизна | большая кривизна | |
| Мясо (150) Масло (75) Масло (75) и через 15 мин мясо (150) | 29,4 2,5 2,6 | 37,4 15,8 37,9 |
Указанной особенностью действия жира па работу желудочных желез отчасти и объясняется своеобразный тип кривой секреции при еде молока, который резко отличается от типа кривых секреции при еде мяса и хлеба. Отсутствие при еде молока резкого подъема кривой в первый час, как это типично для мяса и хлеба, зависит от слабого рефлекторного возбуждения желудочных желез во время еды и угнетающего действия жирных веществ, содержащихся в молоке. Разгар секреции во втором и особенно в третьем часу связан с со-когонным действием образующихся при расщеплении жира мыла, а также продуктов переваривания белков. По мере удаления из желудка пищевых масс секреция постепенно уменьшается и к концу шестого часа вовсе прекращается.
Анализ сокогонного действия различных пищевых веществ позволил дать объяснение кривой секреции при еде мяса и хлеба. Значительное отделение сока при еде; мяса и хлеба в первые 2 ч отражает энергичную работу желудочных желез, вызванную рефлекторно актом еды. Дальнейшее постепенное падение секреторной кривой при приеме мяса и затяжной характер секреторной кривой при приеме хлеба объясняется различным действием химических возбудителей (экстрактивные вещества, вода, продукты переваривания белков), которое вначале суммируется с действием рефлекторных раздражений, благодаря чему наблюдается обильное сокоотделение, а затем, по мере ослабления действия рефлекторных раздражений, возбуждение желез поддерживается по преимуществу влиянием химических раздражений (гормон гастрин, продукты расщепления пищи). Переход пищевых масс в кишечник и ослабление действия химических возбудителей обусловливают снижение и затем полное прекращение желудочной секреции. При еде хлеба первоначальный подъем секреторной кривой связан с рефлекторным возбуждением желез актом еды; последующее скудное отделение сока на протяжении 9 ч объясняется отсутствием в хлебе сильных химических возбудителей желудочных же.чез.
Количество и качество желудочного сока при приеме смешанной пищи бывают различными в зависимости от процентного соотношения входящих в псе основных сортов пищи, а также различных дополнительных веществ, входящих в то или иное блюдо.
Особый интерес представляют данные, полученные II. П. Разенковым и его сотрудниками в опытах па собаках с кормлением их различными блюдами, входящими в рацион питания человека.
Опытами установлено, что при приеме различных супов наибольшее количество желудочного сока отделяется при приеме ячневого, овсяного и картофельного супов и сравнительно меньше — рисового и манного. Значительное количество сока отделяется при еде рассольника и капустных щей, особенно кислых. При приеме молочного киселя, двух яиц и стакана какао секреция желудочного сока длится око.чо 5 ч, отделяющийся при этом сок обладает высокой кислотностью и средней переваривающей силой. Менее обильную секрецию вызывает омлет из 3 яиц и простокваша. Из вторых блюд, в состав которых входят молочные каши, пудинги, суфле из рыбы, вареный судак, наибольшее количество сока отделяется на суфле из рыбы и наименьшее — на рисовый пудинг и манную кашу. Из мясных блюд наибольшее количество сока отделяется при приеме мясного рулета и наименьшее — при приеме макаронника. Большое количество сока выделяется при приеме тушеного мяса и особенно бефстроганов. Из сладких блюд наибольшую секрецию вызывает компот из сухих фруктов с примесью сока сырых апельсинов. Вишневый и молочный кисели являются слабыми возбудителями секреции желудочного сока. При приеме бобовых пищевых веществ, таких, как горох, фасоль, чечевица, бобы, наблюдается секреция сока, во многом напоминающая секрецию сока при приеме хлеба. Если к бобовой пище добавить масло, то наибольшее количество сока бывает при еде чечевицы, а затем гороха, фасоли и бобов. Опытами показано, что объем желудочной секреции, кислотность и переваривающая сила сока при еде молочных каш (манной, рисовой, ячневой) бывают большими, чем при еде тех же каш, но приготовленных на воде. Аналогичные соотношения наблюдаются и при еде пю ре, приготовленных на молоке и воде. К сильным возбудителям секреции желудочного сока относятся бульоны из костей. Их сокогонный эффект превосходит действие либиховского экстракта и мясного бульона. Прибавление крупы к мясному бульону снижает секреторную деятельность желудка по сравнению с действием па желудочные железы одного лишь мясного бульона. При мясном бульоне с фрикадельками наблюдается некоторое замедление процесса сокоотделения по сравнению с секрецией сока при приеме одного мясного бульона.
Детальные исследования были проведены по анализу сокогонного действия различных овощей и их соков. Опытами К. М. Быкова, П. И. Лепорского, М. П. Бре-еткина, И. П. Разепкова и других показано, что пюре из свеклы, моркови, репы и огурцов обладают сильным сокогонным действием. Наибольшее количество сока выделяется на пюре из капусты, огурцов, репы, свеклы и моркови. Наибольшая переваривающая сила бывает при приеме пюре из свеклы и репы и самая низкая — при приеме шоре из моркови. Из фруктов и ягод по со-когонному действию на первом месте стоят персики, затем виноград, апельсины, вишни, груши, чернослив, клубника и, наконец, яблоки. Обычно фрукты вызывают отделение сока с низкой переваривающей силой. Как показали опыты, другие фрукты и ягоды, как, например, черешня, крыжовник, слива, малина и абрикосы, оказывают на желудочные железы тормозящее действие. Вареные и жареные грибы являются сильными возбудителями секреции желудочного сока. По сокогонным свойствам белые грибы и лисички превосходят мясной навар.
По последним данным, в возбуждении деятельности желудочных желез играют роль и витамины (С, В, никотиновая кислота и др.), которые являются отчасти самостоятельными возбудителями секреции, а главным образом активаторами процесса.
При заболеваниях желудочно-кишечного тракта широко применяются с лечебной целью минеральные воды. Систематический прием минеральных вод Кавказских минеральных источников оказывает положительное действие на работу пищеварительных желез, в том числе в желудочных. Установлено, что минеральные воды, принятые с пищей за 5- 15 мин до еды, возбуждают, а принятые за 1 1,5 ч до еды - тормозят деятельность секреторных клеток желудка (рис. 23).
По последним экспериментальным и клиническим наблюдениям механизм действия минеральных вод па желудочные железы нервно-гуморальный. В нем принимают участие рецепторы слизистой рта, пищевода, желудка и кишечника, а также рецепторы стенок кровеносных сосудов, при раздражении которых рефлекторно изменяется деятельность желудочных желез.
При внутреннем употреблении минеральные воды, всасываясь в кишечнике, оказывают гуморальным путем действие как па центральные и периферические ин-нервационпые механизмы, так и непосредственно па нервно-мышечный и нервно-секреторный аппараты желудка.
Многими исследователями было проведено изучение работы желудочных желез при различных пищевых режимах (рис. 24). П. П. Разенков показал, что секреторная реакция желудка при действии тех пли иных раздражителей зависит от функционального состояния желудочных желез, которое может изменяться при нахождении организма длительное время на одном пищевом режиме. Так, если животное долгое время содержится на углеводном режиме, то секреторная реакция желудка характеризуется повышением сокоотделения в первую сложнорефлекторную фазу и понижением количества отделяемого сока во вторую, нервно-химическую фазу; при белковом режиме наблюдаются обратные отношения: уменьшение сокоотделения в первую и увеличение количества сока во вторую фазу секреции. При этом происходит изменение величины латентного периода и качественного состава желудочного сока. Аналогичное явление наблюдается и у людей при длительном пребывании их па одном режиме питания.
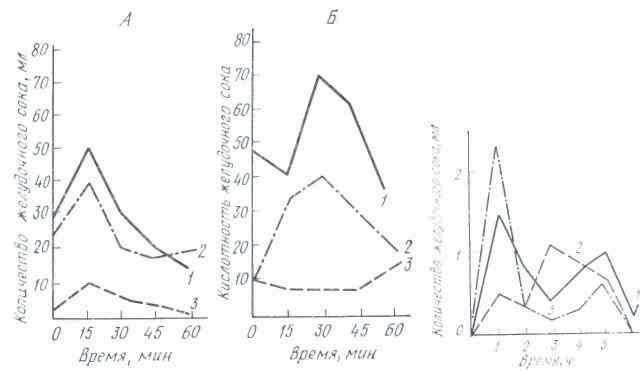
Рис. 23. Влияние однократного приема. железноводекой минеральной воды (Смирновская № 2) на секрецию и кислотность желудочного сока, у человека (по И.Т.Курцину) А — до приема воды, Б — после приема: 2— Кривые сокоотделения hi-контроль; 3—прием минеральной воды за 1 ч до начала исследования.
Рис. 24. Изменение секреции желудочного сока у собаки при длительном нахождении на различном пищевом режиме (по И.П. Разенкову): 1 — при смешан ной пище; 2—при мясном режиме; а – при углеводном режиме
Изменение возбудимости желудочных желез во всех этих случаях обусловлено тем, что длительное пребывание человека пли животного на определенном пищевом режиме изменяет высшую нервную деятельность. Благодаря этому организм приспосабливает работу пищеварительного аппарата и весь процесс пищеварения к различным пищевым режимам, что, несомненно, имеет известное биологическое значение.
Электрические явления в желудке
Деятельность желудочных желез сопровождается особыми ритмическими колебаниями электрического потенциала. Впервые на это явление обратил внимание советский физиолог В.Ю. Чаговец. Он установил тесную связь между секреторной активностью желудочных желез и величиной электрического потенциала (электрогастрограмма).
Последними работами физиологов (А.И. Венчиков и др.) доказано, что и вне пищеварения в железистом аппарате желудка происходит небольшое ритмическое колебание электрического потенциала, по-видимому, связанное с энергетическими процессами, совершающимися в клетках в связи с образованием и накоплением секрета.
Изменение величины потенциала в слизистой желудка при виде нищи или акте еды наступает через 15 — 20 с. после начала действия раздражителя. Амплитуда колебаний электрического потенциала желудка собаки при температуре 20—25° находится в пределах 2 -5 мВ, частота колебаний 2 3 кол/мни. Изменение потенциала происходит в течение всего секреторного периода; возвращение электродвижущей силы к исходной величине наблюдается по прекращении секреции.
Пищевое возбуждение желудочных желез сопровождается усилением эфферентной и афферентной импульсаций в ветвях блуждающих нервов (В.А.Багаев, И.Т.Курцин, А.Д. Ноздрачев). По данным некоторых исследователей, источником возникновения электрических явлений служит не только железистый, но и мышечный аппарат желудка.
С помощью вживленных специальной конструкции электродов у собак в условиях хронического опыта по Павлову удалось записать увеличение афферентной импульсации в желудочных ветвях блуждающих нервов а связи с возникновением и развитием секреторного процесса (В.Л. Багаев, И.Т.Курцин, Л.Д.Ноздрачев). Раздражение мехапорецепторов желудка изменяет электрическую активность в соматосенсорпой области коры головного мозга (С.С.Мусящикова и В. И. Черниговский) (рис. 25).
Структурно-морфологические изменения желез желудка. Исследованиями И. П. Разспкова и его сотрудников установлено, что секреция желудочного сока тесно связана со структурно-морфологическими изменениями секреторных клеток. Так, длительная секреторная деятельность желудочных желез при мнимом кормлении сопровождается усиленным образованием побочных клеток и появлением переходных форм от главных клеток к побочным: обкладочные клетки подвергаются резкой вакуолизации. Усиленное образование побочных клеток является результатом физиологической регенерации же слизистых клеток, так как главные клетки не обладают способностью к регенерации и восстановление их идет за счет передифференцировки побочных клеток.
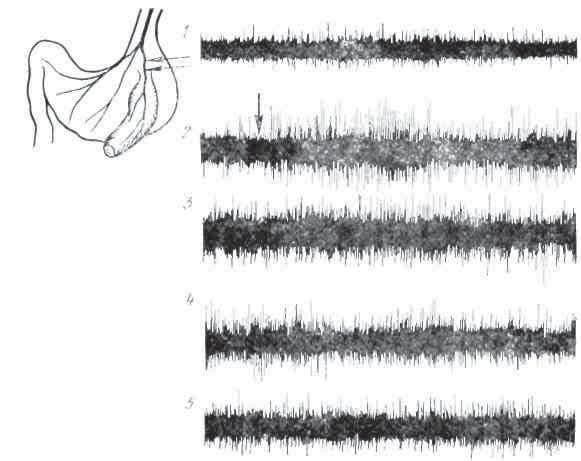
Рис. 25. Усиление афферентной им пульсации в желудочной ветви вагуса в связи с актом еды и начальной фазой желудочного пищеварения у собаки в хроническом опыте: 1 спонтанная осциллограмма; 2 — при акте еды (стрелка); 3 после еды черен 4Г) мин; 4- через 30 мни; 5 — через 3 ч после еды. Слева - схема расположения э лек г родов на веттви вагуса, инннервирующего изолированный по Павлову желудочек
Таким образом, при развитии секреторного процесса и железах желудка происходят структурно-морфологические изменения, которые имеют при нормальных физиологических условиях обратимый характер.
При удалении пилорической части желудка наблюдается отчетливое изменение секреторной функции фундальных желез, сопровождающееся структурно-морфологическим изменением их клеток; желудочный сок в этих случаях содержит значительное количество слизи и обладает большой переваривающей силой. Это происходит благодаря тому, что главные клетки тела желудка превращаются в побочные, которые начинают усиленно проаудировать слизь. Количество побочных клеток возрастает, и из них постепенно формируются пилорические железы. При удалении дна и тела желудка происходят структурно-морфологические изменения в слизистой пи-лорической части, которые выражаются, с одной стороны, в гиперплазии железистого аппарата и, с другой, в атрофии слизистой оболочки, уменьшении объема отдельных пилорических желез; в некоторых зонах привратника железистый аппарат почти полностью исчезает и па его месте разрастается соединительная ткань. В последующем наступает образование новых пплорических желез.
Экспериментальные данные лаборатории И. П. Ра-зенкова о соотношении секреторных и структурно-морфологических изменений в железистых аппаратах желудка при развитии секреторного процесса и после удаления (резекции) различных отделов желудка свидетельствуют о наличии тесной связи между функцией и структурой клеток.
Механизм регуляции секреции желудочного сока относится к числу сложных проблем физиологии, поэтому удобнее провести его анализ раздельно: в первую, слож-норефлекторную, и вторую, нервно-химическую, фазы секреции, хотя эти (разы взаимосвязаны и характеризуют единый секреторный процесс желудка.
Механизм регуляции секреции желудочного сока был изучен благодаря классическим исследованиям 11. П. Павлова и его школы (И. П. Разенков, К. М. Быков, И. С. Цитович, Г. П. Зеленый, В. В. Савич, Л. Л. Орбели, Г. В. Фольборт и его ученики). Существенный вклад в разработку этой проблемы внесли и зарубежные ученые, исследования которых базировались па фундаментальных трудах павловской школы (Айви, Гроссман, Увиес, Бикел, Дрейгштедт, Кач и др.).
Блуждающие нервы, как уже указывалось, являются секреторными нервами желудка. Это впервые было доказано И. П. Павловым и Е. О. Шумовой-Симановской (1889—1890) в хронических опытах на гастроэзофагото-мированных собаках. Они показали, что, во-первых, мнимое кормление вызывает обильное отделение желудочного сока; во-вторых, после перерезки блуждающих нервов это же мнимое кормление не возбуждает желудочных желез и, в-третьих, раздражение периферических концов перерезанных на шее блуждающих нервов электрическим током вызывает секрецию желудочного сока.
В дальнейшем это было подтверждено в многочисленных опытах другими исследователями, причем установили, что блуждающие нервы являются секреторными нервами не только у собак, но и у других живот-пых (кошка, обезьяна, голубь, черепаха, лягушка и др.).
О роли блуждающих нервов как секреторных нервов желудка у человека говорят наблюдения на больных, у которых по медицинским показаниям были наложены фистулы желудка и пищевода. Мнимая еда вызывала у таких людей обильное отделение желудочного сока. У больных, подвергшихся операции перерезки обоих блуждающих нервов, акт еды не возбуждает деятельности желудочных желез. Введение атропина, парализующего периферические окончания блуждающих нервов, также делало невозможным возбуждение желудочной секреции актом еды.
Кроме блуждающих и чревных нервов в секреторной работе желудка принимает участие и правый диафрагмальный нерв. После перерезки его происходит изменение не только моторики, но и секреции, а именно количество сока па пищевые раздражители увеличивается, содержание соляной кислоты и пепсина повышается.
Г.В. Фольборт с сотрудниками экспериментально показали, что чревные нервы принимают участие в механизме возбуждения желудочных желез, поэтому их называют также секреторными нервами. Кроме того, в процессе возбуждения желудочных желез принимают участие мозжечок, гипоталамическая область головного мозга и верхний шейный симпатический узел (Л. А. Орбели и др.). Эти нервные образования подчинены влиянию коры больших полушарий головного мозга.
Согласно данным, полученным в лаборатории К. М. Быкова, регуляция деятельности желудочных желез симпатическими и парасимпатическими нервами построена не по типу антагонизма, как это ошибочно утверждалось некоторыми учеными, а па принципе синергизма, что полностью соответствует точке зрения И. П. Павлова.
Установлено, что у собак с удаленными большими полушариями головного мозга вид и запах пищи не вызывает отделения желудочного сока, а акт еды постоянно дает положительный эффект. Следовательно, за мыкание рефлекторной дуги при безусловном рефлексе с ротовой полости па желудочные железы происходит и подкорковых центрах, путь же условных рефлексов с рецепторов глаза, носа и уха на желудочные железы проходит через кору больших полушарий.
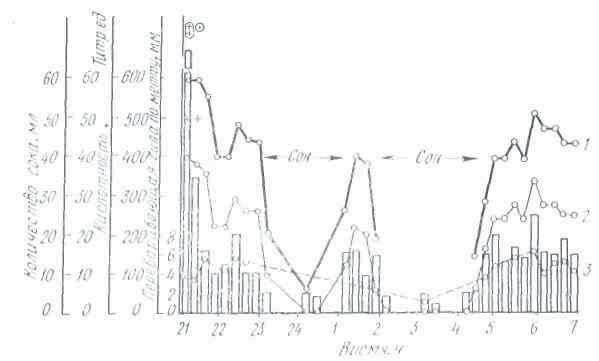
Рис. 26. Ночная желудочная секреции у здорового человека (по Ф. И. Комарову, 1953): 1 — общая; 2 — свободная кислотность; 3 — переваривающая сила (столбики — количество сока, мл). Знак плюс-слизь, плюс и кружке - примесь желчи
У человека во время сна, т. е. когда кора головного мозга находится в состоянии торможения, секреция желудочного сока па пищевые вещества (рис. 2G), введенные в желудок, резко понижена (Ф. И. Комаров).
Сокоотделение в период сложнорефлекторной фазы относится к категории легко тормозимых реакций. Это торможение может быть вызвано импульсами, идущими из нейтральной нервной системы. Различные эмоциональные состояния резко угнетают секреторную деятельность желудка человека.
При виде кошки у собаки наступает агрессивная реакция, сопровождающаяся торможением секреторной функции желудка.
Длительное торможение секреторного процесса возникает у животных при экспериментально вызванном патологическом состоянии клеток коры больших полушарий. Торможение желудочной секреции наблюдается и при раздражении периферических нервов. При сильном раздражении, например, седалищного нерва или рецепторов прямой и слепой кишок, мочевого пузыря и других внутренних органов наступает длительное и резкое угнетение желудочной секреции (Л. А. Нечаев, С.С.Полтырев, А. В. Риккль).
Подобное торможение наблюдается и при болевых раздражениях. Более сильное угнетение деятельности желудочных желез во всех случаях наблюдается на малой кривизне желудка. По данным Л. А. Орбели и его сотрудников, болевые раздражения вызывают торможение желудочной секреции непосредственно через нервные проводники и через инкреторные органы (гипофиз, надпочечники), которые при этом усиленно продуцируют гормоны.
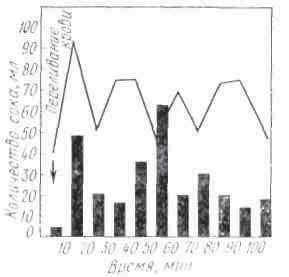 Рис. 27. Секреция желудочного сока у человека после переливания 100 мл крови, взятой от другого человека после мнимого кормления (по П. II. Лепорскому): Столбиками обозначен объем секреции сока. Кривая показывает степень свободное соляной кислоты желудочного сока
Рис. 27. Секреция желудочного сока у человека после переливания 100 мл крови, взятой от другого человека после мнимого кормления (по П. II. Лепорскому): Столбиками обозначен объем секреции сока. Кривая показывает степень свободное соляной кислоты желудочного сока
В последнее время физиологами установлено, что в слизистой оболочке тонкой кишки образуются специфические вещества, которые тормозят желудочную секрецию гуморальным путем. Эти вещества по химическому свойству тождественны гормону энтерогастрону, выделенному в чистом виде из слизистой кишечника и действующему тормозящим образом на желудочные железы. Аналогичное действие оказывает на них и гормон урогастрон, полученный из мочи человека и животных. Этот гормон в целостном организме оказывает тормозное действие на секреторные клетки желудка при участии нервных элементов, что соответствует положению И. П. Павлова о единстве и взаимосвязи нервного и гуморального механизмов регуляции.
В процессе возбуждения и торможения секреторных клеток желудка принимают участие и химические передатчики нервного, возбуждения (медиаторы). По характеру действия они могут быть ацетилхолиноподобными или адреналиноподобными веществами. Первые оказывают на желудочные клетки возбуждающее, а вторые — тормозящее действие. И. П. Разенков обнаружил в крови собаки в момент раздражения периферического кон-па блуждающего нерва биологически активные вещества, которые при введении в кровь другой собаки вызывали секреторный эффект. Ряд исследователей обнаружили медиаторы в крови, оттекающей от желудка, а также в стенке желудка и желудочном соке. В начале желудочной секреции, вызванной показом пищи животному или мнимым кормлением, в крови резко увеличивается количество ацетилхолиноподобных веществ (В. Г. Прокопенко). В специальных исследованиях на людях Н. И. Лепорский и Е. А. Нечаева показали,что кровь одного человека, взятая в разгар секреции, вызванной натуральными условными раздражителями, и перелитая другому человеку, обладает свойством возбуждать у последнего желудочную секрецию (рис. 27). Возможно, что усиленное образование ацетилхолиноподобных веществ в этих случаях происходит преимущественно в центральных инпервационных приборах, хотя не исключено и образование их на периферии, в частности в желудке.
Регуляция секреторного процесса обусловливается пищевым центром, расположенным в различных отделах головного мозга.
Высший регуляторный орган — кора больших полушарий, которая оказывает влияние на работу желудочных желез по типу «пускового» и «корригирующего» механизмов, т. е. вызывая активность желез при их физиологическом покое («пусковой» механизм влияний).
В возникновении секреторной реакции большую роль играет исходная возбудимость желудочных желез, которая определяется функциональным состоянием пищевого центра.
Передача возбуждения с рецепторных полей на желудочные железы осуществляется рефлекторным путем по нервам при участии медиаторов (аце-тилхолина и адреналина). Секреторный процесс возникает вначале в железах малой кривизны, а затем в железах других секреторных полей желудка. Величина латентного периода возбуждения желудочных желез на малой кривизне равняется 1—3 мин, а на большой кривизне — 5—10 мин, если судить по первой кислой капле, выделившейся через фистулу наружу. На самом деле, как показали последние электрофизиологические исследования, он исчисляется от 7 до 20 с,
Рефлекторное возбуждение желудочных желез легко тормозится при воздействиях на высшие отделы центральной нервной системы и при раздражениях периферических нервов. В механизме торможения секреторного процесса принимают участие и инкреты желез внутренней секреции (адреналин и питуитрин), а также специфические гормоны, образующиеся в стенке кишечника (энтерогастрон); тормозящее действие их на секреторные клетки осуществляется через нервную систему.
Желудочная секреция, возникающая при механическом раздражении рецепторов желудка, — сложнорефлекторная реакция, в которой принимает участие кора больших полушарий головного мозга. «Механическая секреция» возникает вначале в секреторном поле малой кривизны, а затем в других секреторных полях желудка. У человека она характеризуется коротким, по сравнению с животными, латентным периодом и обильным отделением сока в первые 15—30 мин непрерывного действия раздражителя. Количество отделяемого сока зависит, с одной стороны, от функционального состояния высших отделов центральной нервной системы и степени возбудимости нервно-железистого аппарата желудка, с другой — от силы и времени действия раздражителя.
Возбуждение фундальных желез при механическом раздражении стенок дна и тела желудка осуществляется при помощи рефлекторного механизма. Рефлекторная передача возбуждения с механорецепторов желудка на секреторные клетки происходит через блуждающие нервы.
В процессе возбуждения желудочных желез человека большую роль играет кора головного мозга — при ее участии происходит образование условного интероцеп-тивного рефлекса. Поскольку у людей во время сна «механическая секреция» бывает меньше, чем во время бодрствования, то, следовательно, механизм возникновения ее у человека осуществляется помимо условного интероцептивного рефлекса и в порядке безусловнореф-лекторной реакции (рис. 28).
Нервно-химическая фаза желудочной секреции возникает вслед за сложнорефлекторной и обусловлена возбуждающим действием специфических гормонов, продуктов расщепления пищевых веществ и экстрактивных, содержащихся в мясе, рыбе и овощах и появляющихся при варке супов. Основанием для признания наличия второй фазы желудочной секреции послужили опыты на собаках с частично (по Гейденгайну) или полностью (по Грегори) денервированными изолированными желудочками. Под влиянием вводимых в полость большого желудка экстрактивных веществ в изолированном желудочке начиналось отделение кислого желудочного сока. В то же время акт еды подобной реакции не вызывал. Изучение механизма развития секреторного процесса в эту фазу дало много ценных сведений. Однако до сих пор механизм работы желудочных желез еще недостаточно ясен и по этому поводу существует несколько теоретических воззрений.
Теория И. П. Павлова
И. П. Павлов утверждал, что «система организма, его бесчисленных частей соединяется в единое целое двояким образом: посредством специфической ткани, которая существует только для поддержания взаимных отношений, а именно нервной ткани, и при помощи тканевых жидкостей, омывающих все тканевые элементы. Эти же самые посредники переносят также и наши раздражители на железистую ткань» Павлов И. П. Поли. собр. соч., т. II, кн. 2. М., 1951, с. 355—356.
Механизм второй фазы желудочной секреции И. П. Павлов рассматривал как нервно-химическую. Об этом свидетельствует ряд фактов. Например, введение атропина в разгаре пищеварения полностью прекращает секрецию желудочного сока. Подобный эффект наблюдается и в тех случаях, когда перед введением химических возбудителей в привратник производилась предварительная кокаинизация слизистой оболочки последнего. Установлено также, что у людей и животных после двусторонней перерезки блуждающих нервов значительно уменьшается секреторный эффект на химические возбудители и т. д.
Теория И. С. Эдкинса
Ряд физиологов, стоявших на позициях «гуморализма», рассматривает вторую фазу желудочной секреции как чисто гуморальную, не связанную с нервной системой. Такого же взгляда придерживается американский ученый Эдкинс, выдвинувший теорию гормонального возбуждения желудочных желез. Согласно этой теории возбуждение фундальных желез во второй фазе обусловливается действием специального гормона гастрина, который под влиянием химических веществ образуется в стенке пилорического отдела желудка из прогормона и кровяным током подносится к фундальным железам. В опытах было показано, что кровь, взятая у собаки с удаленным привратником в разгаре пищеварения и перелитая другой собаке, не вызывает секреции желудочного сока, в то время как кровь собаки с сохраненным пилорическим отделом желудка всегда вызывает секрецию. Ряд авторов утверждает, что в привратнике образуются гормоны иило-рин и пилороантрумгастрин, которые гуморальным путем возбуждают фундальные железы.
Теория Л. Б. Попельского
Теория гормонального механизма возбуждения желудочных желез получила дальнейшее развитие в исследованиях Л. Б. Попельского, установившего, что в процессе возбуждения желез принимает активное участие гистамип, образующийся в желудке. Является ли гистамин действующим началом гастрина Эдкинса, как это считал По-пельский и другие исследователи, или он представляет самостоятельный гормон в настоящее время точно не установлено. По мнению некоторых авторов, гистамин никакого отношения к гастрину не имеет. Возможно, гистамин— медиатор, выделяющийся при раздражении блуждающих нервов.
Эндокринные теории
Кроме гастрина, пилорина, пилороантрумгастрина и гистамина в последнее время были обнаружены в слизистой оболочке верхних и средних отделов тонкого кишечника собак особые гормоны, обладающие способностью возбуждать желудочную секрецию («желудочные секретины»). Помимо названных гормонов, в желудке был недавно найден еще один гормон, обозначенный субстанцией «Р», который также обладает свойством возбуждать желудочные клетки. В механизме желудочной секреции принимают участие и такие гормоны желез внутренней секреции, как инсулин, адреналин, паратиреоидин и гидрокортизон. Особая роль в этом отношении принадлежит инсулину, который возбуждает желудочные железы через систему блуждающих нервов, и адренокортикотропному гормону (АКТГ), который стимулирует образование соляной кислоты желудочного сока через кортикостероид-ные гормоны надпочечника.
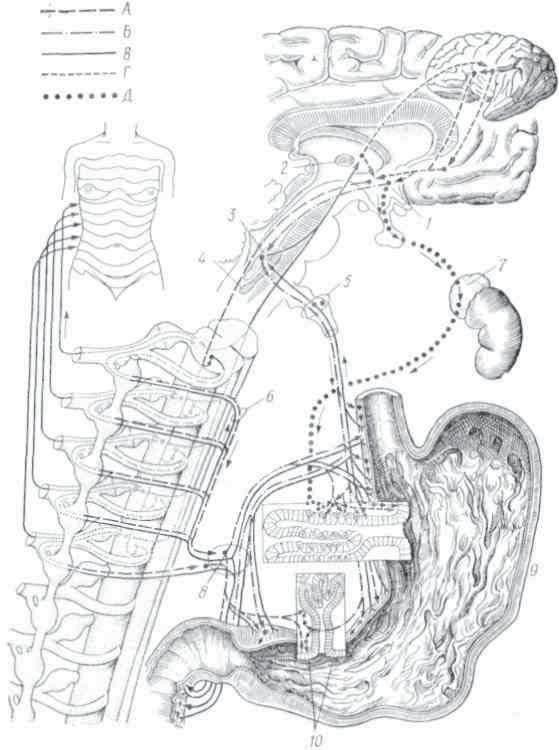
Рис. 28. Нейрогуморальная регуляция функций желудка (по Неттер, 1959).
А — симпатические эфферентные волокна, Б — парасимпатические эфферентные волокна, В — афферентные волокна, Г— неизвестные пути, Д — гуморальные связи: 1 — гипоталамус; 2 — таламус; 3 — дорзальпые ядра вагуса; 4 — tr. solita-rius; 5 — блуждающий нерв; 6 — чревный нерв; 7 — кора надпочечников; 8— gangl. coeliacum; 9 — фуидальные железы; 10 — пилорические железы
Теория С. Окада
Участие инсулина и адреналина в регуляции концентрации сахара крови дало основание японскому ученому Окада и некоторым другим исследователям высказать теорию о механизме регуляции деятельности желудочных желез. Согласно этой теории при повышении уровня сахара крови (гипергликемия) угнетаются, а при понижении уровня сахара крови (гипогликемия) возбуждаются секреторные клетки желудка. Однако эта теория встретила серьезные возражения. Вероятно, участие сахара крови в процессе возбуждения желудочных желез следует рассматривать в связи с общей нервно-гуморальной регуляцией, осуществляемой пищевым центром.
Теория И.П. Разенкова
И.П. Разенков в 1925 г. выдвинул теорию чисто химического возбуждения желудочных желез. Согласно ей механизм второй Фазы желудочной секреции не гуморальный, с точки зрения специфического гормона, а чисто химический, поскольку химические возбудители, т. е. продукты переваривания пищи, попав в круг кровообращения, непосредственно сами возбуждают работу желудочных желез. Проведенный анализ «сытой» и «голодной» крови показал существенную разницу биохимических свойств той и другой. На собаках с удаленным привратником было установлено, что пищевые и химические вещества (молоко, либиховский экстракт, гемоза, соляная кислота и др.), введенные в тонкую кишку, возбуждают деятельность желудочных желез.
Подобный эффект наблюдается и у больных с удаленным привратником.
В 1948 г. И. П. Разенков пришел к заключению, что вторая фаза секреторной деятельности желудочных желез обусловливается не только действием составных частей пищи и продуктов ее переваривания, но и специфически гормональными веществами, образующимися в слизистой оболочке пилорической части желудка или кишечника.
Итак, вторая фаза желудочной секреции является очень сложной; в ней принимают участие, помимо рефлекторных рлияний различных веществ с привратника и двенадцатиперстной кишки, специфические вещества гормонального происхождения (гастрин, пилорин, пило-роантрумгастрин, гистамин, субстанция «Р» и т. д.) и продукты переваривания самой пищи, которые через кровь поступают в желудок и возбуждают его секреторные клетки.
Объяснить механизм возбуждения желудочных желез во вторую фазу секреции какой-либо одной из предложенных гуморальных теорий, с точки зрения современных представлений, основанных на принципах павловского «нервизма», невозможно.
Установленные И. П. Павловым и его сотрудниками, а позднее А. В. Соловьевым, Е. М. Матросовой, А. Н. Ба-курадзе факты о зависимости работы желудочных желез во вторую фазу секреции от нервной системы (симпатической и парасимпатической) дают право считать, что в механизме возбуждения желудочных желез гормонами и химическими веществами принимает участие нервная система. Если еще принять во внимание, что в механизме возбуждения желудочных желез во второй фазе их деятельности принимают большое участие и медиаторы, то станет вполне очевидным, что механизм возбуждения желез желудка во вторую фазу является нервно-гуморальным. Такой взгляд полностью отражает принципиальное положение И.П.Павлова о единстве нервного и гуморального механизмов регуляции при ведущем значении нервного механизма.
Двигательная деятельность желудка
Гладкие мышцы в отличие от поперечнополосатых не имеют выраженной исчерченности, сокращаются медленно, отвечают сокращением на растяжение и могут находиться в сокращенном состоянии продолжительное время без признаков утомления. По степени зависимости от внешней иннервация и способности реагировать па раздражение их можно разделить на две группы: 1) мышцы, которые сокращаются с определенной силой только в ответ на возбуждение их внешних нервов и не обладают или почти не обладают спонтанной активностью, и 2) мышцы, характеризующиеся способностью к спонтанной ритмической активности, которая изменяется под влиянием пусковых нервов, но определяется не нервами, а свойствами мышечных клеток (мышцы пищеварительного тракта).
Сократительная деятельность гладких мышц пищеварительного тракта регулируется экстрамуральными и интрамуральными нервными, миогенными, гуморальными и сосудистыми механизмами.
Клетка гладкой мышцы имеет веретенообразную форму с ядром в средней, самой широкой части. Длина клеток чаще колеблется от 100 до 200 мкм, диаметр — 2—10 мкм, толщина циркулярного слоя мышц — 5 мкм. При сокращении длина и форма клеток изменяются. Каждая клетка имеет плазматическую мембрану, которая снаружи покрыта базальной мембраной. Обе мембраны составляют сарколемму — поверхностную мембрану. Ба-зальная мембрана отделена от плазматической промежутком в 6—13 нм. На поверхности мембраны клеток гладких мышц много вдавлений внутрь клетки в виде пузырьков— пиноцитов. Они составляют часть функциональной структуры клетки, которая имеет важное значение для проницаемости клеток и деятельности гладких мышц. Общая поверхность клетки, не учитывая пиноци-ты, составляет около 400 мк2. На поверхности каждой клетки находится около 100—150 отростков, обеспечивающих боковые мембранно-мембранные контакты с соседними клетками. Отростки увеличивают поверхность клетки, во время мышечного сокращения они втягиваются внутрь клетки, одновременно увеличивается также количество и размеры пиноцитных пузырьков.
Внутреннее содержимое клетки — основное бесструктурное вещество (саркоплазма, гиалоплазма), в котором расположены саркоплазматическая и эндоплазматические сети. Она участвует в осуществлении связи процессов на поверхностной мембране с сократительным аппаратом. В саркоплазме имеются ультраструктурные образования — сократительный протофибриллярный аппарат. Сократительный аппарат состоит из про-тофибрилл, сгруппированных в нити — миофибриллы, которые размещаются в клетке параллельно друг другу.
При помощи отростков и сети коллагеновых волокон гладкомышечные клетки группируются в длинные пучки, являющиеся функционально эффекторными единицами в мышечных слоях стенки желудочно-кишечного тракта, они разделены соединительноткаными перегородками. Диаметр пучков около 100 мкм. Они ветвятся и при этом создают тяжи переходов от одного пучка к другому, что важно для деятельности мышцы как единой системы.
По вопросу иннервации клеток гладких мышц и связях с ними постганглионарных парасимпатических и симпатических аксонов, о роли нервных сплетений, интерстициальных клеток и нервной сети нет единого мнения. Известно, что окончания нервных волокон никогда не проникают внутрь плазматической мембраны. Они располагаются в своеобразных выемках клеток гладких мышц в виде расширенного окончания или проходят вблизи клеток, контактируя с ними. Передача нервных влияний на гладкомышечные клетки осуществляется посредством медиаторов нервного возбуждения — ацетилхолина, норадреналина.
Распределение ионов между клетками гладких мышц и внеклеточной окружающей средой иное, чем в скелетных мышцах. В гладких мышцах более высокое содержание хлористого натрия и большая проницаемость клеток к некоторым ионам и воде. Неодинаковая проницаемость ионов через полупроницаемую мембрану и работа насоса, выталкивающего ионы из клетки (Na+) или захватывающего их из внешней среды (К+), приводят к асимметричному распределению ионов по обе стороны мембраны клетки, создающему определенную величину трансмембранного потенциала. Главным определяющим фактором мембранного потенциала являются ионы К+, которые обладают высокой проницаемостью через мембрану и содержатся в клетке в большей концентрации, чем в жидкости внеклеточного пространства.
Возбуждение гладкомышечной клетки может распространяться на соседние клетки и на значительные расстояния по гладкомышечной ткани. Это касается медленных волн и потенциала действия. Медленные волны распространяются с большой скоростью, они передвигаются в кишке по продольному слою мышц на расстояние 80—130 см. Распространение потенциала действия ограничено от 0,5 до 3—5 мм и происходит по ходу пучков гладкомышечных клеток.
Передача возбуждения в гладких мышцах пищеварительного тракта от клетки к клетке происходит при помощи электрического тока потенциала действия при участии медленных волн. Для понимания механизмов связи процессов возбуждения и сокращения необходимы данные о структуре и свойствах белков, осуществляющих процесс сокращения. К белкам сократительного механизма относятся актин и миозин. Взаимодействие их приводит к образованию актомиозина, нити которого при действии определенных факторов могут сокращаться. Протофибриллы состоят из актина. Миозино-вых нитей в гладких мышцах не выявлено. Свойства актомиозина, полученного из мышц пищеварительного тракта, отличаются от свойств актомиозина скелетных мышц по Са-активируемой АТФазной активности, инфракрасным спектрам поглощения и ультрафиолетовой люминесценции (П. Г. Богач).
Для сократительного процесса необходима энергия, которая поставляется макроэргическими молекулами, в особенности АТФ и креатинфосфата. Образование реактивного миозинфосфатного комплекса является обязательным условием при взаимодействии миозина с АТФ и важной реакции мышечного сокращения. Известно, что гладкие мышцы потребляют относительно небольшие количества кислорода. При раздражении электрическим током или ацетилхолином напряжение мышцы и потребление кислорода увеличивается, а при действии атропина или адреналина, снижающих напряжение мышцы, потребление кислорода уменьшается. Теплопродукция гладкой мускулатуры прямо пропорциональна потреблению кислорода. Обогащение глад-комышечной ткани кислородом приводит к усилению спонтанной активности, повышению возбудимости и увеличению сопротивляемости мембраны. Добавление С02, а также замена кислорода в газовой смеси азота ведут к противоположным результатам (М. Ф. Шуба).
Желудок выполняет важную двигательную функцию, обеспечивающую, во-первых, превращение пищи в химус в пилорическом отделе и, во-вторых, эвакуацию желудочного содержимого в двенадцитиперстную кишку.
Методика исследования двигательной функции желудка. Двигательная функция желудка изучается при помощи различных методов: манометрографического, рентгенографического, эвакуаторного, гастротонометрического и электрогастрографического.
Наиболее распространенным в физиологии является баллоноки-мографический метод. Он состоит в том, что животному через рот или фистулу вводится в желудок резиновый баллон, который затем наполняется воздухом или водой. При помощи резиновых трубок баллон через водяной манометр соединяется с капсулой Марея. Наступающие вследствие сокращений стенок желудка изменения давления в баллончике передаются капсуле, рычажок которой записывает ни ленте кимографа кривую сокращений.
У человека можно записывать сокращения желудка и одновременно исследовать секрецию желудочного сока.
В клинике особенно распространен рентгенографический метод, при котором больной съедает кашу с примесью нерастворимых солей бария, обладающих способностью задерживать рентгеновские лучи. При просвечивании на экране получаются отчетливо выраженные контуры желудка, а последовательный ряд снимков дает представление об изменениях его формы и величины, о скорости эвакуации введенной массы в двенадцатиперстную кишку, функциональном состоянии пилорического сфинктера. Следует отметить, что со времени применения впервые Кенноном рентгенологического метода для исследования движений пищеварительного тракта (1898) он претерпел дальнейшее совершенствование. В настоящее время осуществлен ряд модификаций этого метода, некоторые из которых позволяют получать более обширную информацию о состоянии двигательной функции желудочно-кишечного тракта. В частности, цепной является рентгенокинематографическая методика, однако обработка получаемых с ее помощью данных трудоемка. К тому же исследуемый получает большую дозу лучей Рентгена. Возможно, с развитием скоростной киносъемки и при использовании сверхчувствительных пленок опасность применения этой методики будет меньше. С. Д. Гроп-сман и А. Д. Беген предложили объемнометрическую рентгеноскопию, позволяющую исследовать динамику эвакуации содержимого фун-дальпого отдела желудка.
Существенное дополнение к рентгенологическому методу — запись сокращений желудка с помощью электрогастрографа (М.Л.Собакин) и радиотелеметрический метод (А. С. Белоусов). Радиотелеметрический метод позволяет одновременно получать сведения о двигательной активности желудка и кислотности сока. Проглатываемая человеком радиокапсула имеет два датчика — давления и рН.
Эвакуаторный метод состоит в том, что в желудок вводится известное количество пищи и затем определяется срок ее эвакуации в двенадцатиперстную кишку. У человека определение скорости эвакуации производится или по изменению интенсивности окраски химических веществ (фенолфталеин, метиленовая синь и др.), добавленных к пробному завтраку, или по величине остатка пробного завтрака, который откачивают через некоторый промежуток времени, или по импульсам, полученным от проглоченной радиокапсулы. Некоторые клиницисты определяют скорость эвакуации по степени кислотности отдельных порций желудочного содержимого, извлекаемого из желудка через равные промежутки времени после введения пробного завтрака. В этих случаях расчет производится по специальным математическим формулам. Для изучения эзакуаторной функции желудка животным накладываются хронические фистулы желудка и двенадцатиперстной кишки. Пища или испытуемые растворы вводятся в желудок через фистулу. По количеству выделившейся за определенное время из фистулы двенадцатиперстной кишки пищи или раствора судят об эвакуации пищи из желудка в кишечник.
При различных заболеваниях сокращения желудка могу-i усиливаться (гиперкинез) или ослабляться (гипокинез), а в ряде случаев и совершенно исчезать (акинез). Например, при язвенной болезни они бывают в большинстве случаев резко выраженными. Перистальтические сокращения возникают в области кардии и волнообразно распространяются до привратника. Благодаря этим сокращениям происходит перемещение пищевых масс по направлению к двенадцатиперстной кишке. Ритмические сокращения способствуют, главным образом перемешиванию желудочного содержимого преимущественно в пилорическом отделе желудка.
Мышечная стенка желудка находится постоянно в состоянии определенного тонического сокращения, которое обусловливает перистолу, возникающую в препилорической части желудка и разделяющую последний как бы на две части. Величина тонического сокращения фундальной части пустого (голодного) желудка может быть различной в зависимости от многих внешних и внутренних причин. У здорового человека она колеблется в пределах от 2 до 1G см вод. ст. Сила отдельных сокращений пилорического отдела желудка у человека достигает 20—70 см вод. сг.
Прием пищи изменяет моторику пилорического отдела желудка: сокращения становятся более сильными, а тонические — более длительными. Эти изменения обусловлены влиянием импульсов, поступающих из верхних отделов центральной нервной системы, с рецеп-торных полей желудка и двенадцатиперстной кишки.
Движения желудка при поступлении пищи
При поступлении пищи в желудок «голодная периодика» сменяется непрерывными сокращениями, продолжающимися в течение всего пищеварительного периода. Эти сокращения связаны с действием на рецепторы желудка механических и химических раздражителей.
Адекватным возбудителем и усилителем моторной деятельности желудка человека и животных является механическое раздражение, оказываемое пищевой массой на рецепторы, заложенные в стенках желудка.
Механизм эвакуации содержимого желудка в двенадцатиперстную кишку в настоящее время недостаточно изучен.
В регуляции движений пилорического сфинктера играют роль химические (соляная кислота, жир, соли и др.) и механические раздражения слизистой двенадцатиперстной кишки. Согласно теории Кеннона, открытие пилорического сфинктера происходит вследствие раздражения рецепторов слизистой выходной части желудка соляной кислотой желудочного сока. При этом часть кислой пищевой массы эвакуируется в двенадцатиперстную кишку. Кислота, действуя на рецепторы слизистой кишки, вызывает рефлекторным путем закрытие пилорического сфинктера и, следовательно, прекращение эвакуации. Рефлекторное закрытие пилорического сфинктера называется запирательным пилорическим рефлексом. После нейтрализации в двенадцатиперстной кишке кислого содержимого щелочным дуоденальным соком реакция опять становится щелочной и пилориче-ский сфинктер вновь открывается. Такая «игра сфинктера» повторяется до тех пор, пока вся пищевая масса не эвакуируется из желудка в кишечник.
Необходимо отметить, что теория Кеннона в последние годы уже не является общепризнанной. Столь сложный процесс, каким является эвакуация содержимого из желудка в двенадцатиперстную кишку, сводить к кислотному рефлексу невозможно. Кстати, еще сотрудниками И. П. Павлова В. Н. Болдыревым, П. Л. Арбековым и Я. Г. Эдельманом было показано, что пилорус не имеет такого исключительного значения в эвакуации из желудка, какое ему первоначально приписывали. Переход содержимого зависит как от сокращения стенок желудка, так и от моторики двенадцатиперстной кишки.
Г.Ф. Коротько выдвинута концепция, согласно которой время пребывания химуса в желудке определяется не только степенью разжижения его, но и соотношением между количеством гидролитически активного секрета, субстратом и продуктами гидролиза в его полости. Вместе с тем в настоящее время в желудке еще не обнаружены рецепторы, реакция которых на химический состав пищи существенно отражалась бы на его моторной деятельности. В желудке были обнаружены рецепторы, реагирующие на рН желудочного содержимого, что указывает на возможное участие кислотности сока в осуществлении эвакуации.
Данные исследований П.Г. Богача дают основание признать, что скорость эвакуации из желудка зависит от объема и степени измельчения желудочного содержимого.
Механизм сокращений желудка. Сокращения желудка обусловливаются заложенным в его стенках нервным аппаратом (ауэрбахово сплетение), который особенно развит в области малой кривизны. Поэтому желудок, изолированный и лишенный связи с центральной нервной системой, способен ритмически сокращаться. Следует, однако, отметить, что, по мнению большинства современных исследователей, ритмические сокращения желудка миогенной природы. Некоторые ученые придерживаются противоположной точки зрения, считая, что они нейрогенного происхождения.
В здоровом организме регуляция движений желудка осуществляется центральной нервной системой, причем с блуждающим нервом к желудку идут стимулирующие, а по чревным — преимущественно тормозящие импульсы.
Двусторонняя перерезка блуждающих нервов сопровождается у собак фазными измененими моторики желудка. Первоначально возникает резкое ослабление сокращений, затем появляются непрерывные сильные сокращения редкого ритма и сократительная активность желудка стабилизируется. У одних собак фаза стабилизации характеризуется стойким ослаблением моторики или полным отсутствием периодов работы, у других— наблюдается тенденция к нормализации пищевой и периодической моторики.
Двусторонняя перерезка чревных нервов вызывает менее существенные изменения моторики желудка, чем ваготомия (С. Д. Гройсман).
Атропин угнетает движения желудка и снижает его тонус как у человека, так и у животных. Аналогичное действие в большинстве случаев оказывает и адреналин. Введение в организм эрготоксина или пилокарпина вы-з.ывает повышение тонуса, учащение и усиление перистальтики желудка.
По последним данным, в регуляции движений желудка принимает участие правый диафрагмальный нерв, раздражение которого ведет то к усилению, то к торможению сокращений желудка.
На моторику желудка также оказывает влияние гуморальным путем ряд неорганических и органических веществ (гистамин, холин, адреналин, продукты расщепления белка, витамины, растворы кальция, сахара и т. д.). Так, гистамин, холин и его производные (аце-тилхолин) резко усиливают сокращения желудка; некоторые считают эти вещества специфическими гормонами, возбуждающими моторику желудка.
Двигательная реакция желудка на механическое раздражение осуществляется при помощи двоякого рода механизмов: местного, интрамурального, и общего, сложнорефлекторного. Последний у человека и высших животных подчиняет себе деятельность местного интрамурального механизма.
В осуществлении двигательной реакции желудка па механическое раздражение принимают участие как механорецепторы слизистой, так и механорецепторы глубоких слоев стенок желудка.
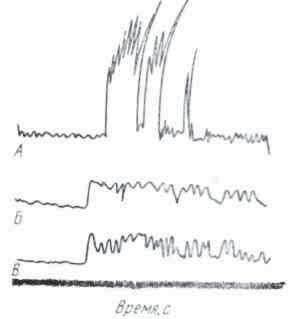
Рис. 29. Изменение моторики желудка и кишечника у собаки при рвоте (по Е.Б. Бабскому).
А — движение желудка, Б — двенадцатиперстнойкишки, В — тонкой кишки.
Тонус мускулатуры желудка и перистальтические сокращения резко изменяются под влиянием импульсов, приходящих из высших отделов центральной нервной системы. Кора больших полушарий головного мозга играет важную роль в регуляции двигательной функции желудка. Свое влияние на желудок она осуществляет при помощи вегетативных нервов и желез внутренней секреции (гипофиз, щитовидная железа, надпочечники и др.
Рвота. Благодаря двигательной функции желудка осуществляется акт рвоты, возникающий рефлекторно при раздражениях, действующих на чувствительные окончания нервов, или автоматически, т. е. через кровь прямо на рвотный центр. Акт рвоты может возникнуть, -например, при раздражениях рецепторов корня языка, носоглотки, слизистой пищевода и желудка, тонкой и толстой кишок, брюшины, органов мочеполовой, дыхательной и других систем организма. Иногда рвота поникает при раздражении обонятельных и вкусовых рецепторов. Автоматически рвота возникает при отравлении ядами, введенными извне (апоморфин и др.), или образовавшимися в организме как при физиологических (беременность), так и патологических (воспаление, сепсис и т. п.) условиях.
Рвота — сложнорефлекторный,координированный двигательный акт защитного характера. Обычно он начинается с антиперистальтики кишечника, в результате чего его содержимое поступает обратно в желудок, а спустя 10—20 с происходит раскрытие кардиального сфинктера, резкое сокращение мускулатуры желудка при сильном напряжении мышц брюшного пресса и диафрагмы. Все это ведет к выбрасыванию содержимого из желудка через пищевод и полость рта наружу (рис. 29). При рвоте происходит рефлекторное сокращение мышц мягкого нёба и оттягивание корня языка книзу, благодаря чему закрывается вход в полость носа и гортань, что предотвращает попадание рвотных масс в носовую полость и дыхательное горло.
Афферентными нервами, участвующими в акте рвоты, являются блуждающие, языкоглоточные и другие нервы эфферентными — блуждающие и симпатические нервы; иннервирующие кишечник, желудок и пищевод, а также ветви, иннервирующие диафрагму и брюшную стенку. Рвотный центр находится в продолговатом мозге, на дне четвертого желудочка. Возбуждение этого центра можно вызвать в порядке условного рефлекса, если какой-либо индифферентный агент сочетать во времени с действием раздражителя, вызывающего акт рвоты (В.Л. Крылов, Н.А. Подкопаев). Рвотные движения возникают и при сильных сотрясениях головы. В этих случаях происходит непосредственное раздражение рвотного центра.
Особенности желудочного пищеварения у разных видов сельскохозяйственных животных
Желудочное пищеварение у жвачных. Анатомическое строение желудка и процесс пищеварения у жвачных животных своеобразны.
Желудок жвачных животных разделяется на четыре отдела: рубец, сетку, книжку и сычуг (рис. 30). Рубец коров, например, представляет собой мышечный мешок, емкость которого достигает 200 л.
Проглоченный пищевой комок попадает в преддверие рубца, а затем в рубец, из которого через некоторое время вновь возвращается в ротовую полость для повторной механической переработки и тщательного смачивания слюной. Этот процесс называется жвачкой. Переход пищевых масс из рубца в ротовую полость осуществляется по типу рвотного акта, в основе которого лежит рефлекторный механизм. Среди эфферентных нервов, принимающих участие в отрыгиваиии, важная роль принадлежит блуждающему нерву, иннервирую-щему преджелудки жвачных. При перерезке блуждающих нервов жвачка прекращается.
Отрыгивание жвачки происходит в результате взаимосвязанных координированных процессов. Последовательно сокращаются сетка и диафрагма, при этом замыкается гортань и открывается кардиальный сфинктер пищевода.
Обычно жвачка начинается через 30—70 мин после еды и протекает в определенном для каждого вида животных ритме.
Продолжительность пребывания каждой порции жвачки во рту — около одной минуты. Поступление следующей порции в рот происходит через 3— 10 с. Продолжительность периода жвачки равна 45— 50 мин, после чего наступает период покоя, длящийся у различных животных разное время, вслед за которым вновь наступает период жвачки. В течение дня корова пережевывает таким образом около 60 кг содержимого рубца.
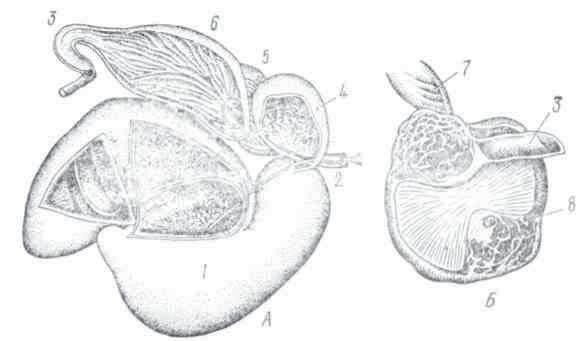
Рис. 30. Схема сложного желудка жвачных животных (А) и желудка птиц (Б):
1 — рубец; 2 — конец пищевода; 3 — двенадцатиперстная кишка; 4 — сетка 5 —книжка; 6 — сычуг; 7 — железистый желудок; 8—мускульный желудок
Пища, подвергшаяся жвачке, затем повторно проглатывается и поступает в рубец и в нем смешивается со всей массой рубцового содержимого. Перемешивание пищи и продвижение ее от преддверия рубца к сычугу происходит благодаря мощным сокращениям мышц преджелудков.
Пищеводный желоб и сетка сокращаются одновременно. Сокращения сетки происходят в две фазы. В первую фазу сетка уменьшается в размерах почти на 50%, згтем слегка расслабляется, после чего вновь наступает сильное сокращение. При второй фазе полость сетки резко уменьшается, и в это время содержимое ее выталкивается в преддверие рубца и частично в книжку.
Вслед за сокращением пищеводного желоба и сетки происходит сокращение рубца. Одна часть сжимается и выдавливает содержимое в другую, расслабленную часть. После этого сокращается следующая часть рубца. Таким образом происходит передвижение и перемешивание пищи в рубце.
Во время сокращения сетки расширяется сычуг, и в нем создается отрицательное давление, в результате которого жидкая масса насасывается из книжки в сычуг, а из сетки в книжку. Обратно пища из книжки в сетку поступить не может, так как этому препятствует сфинктер, находящийся между этими отделами. Так пищевая масса поступает в сычуг, где, собственно, и происходит ее переваривание сычужным соком, выделяемым железами, заложенными в слизистой оболочке. Сок имеет кислую реакцию и содержит фермент пепсин. В принципе процесс, происходящий в полости сычуга, мало чем отличается от процесса переваривания пищи в желудке собаки. Следует отметить, что в слизистых оболочках других отделов желудка жвачных пищеварительные железы отсутствуют.
Кислотность сычужного сока равна 0,043%, доходя у некоторых жвачных до 0,25% (коза). Переваривающая сила сока 5,5 мм (по Метту).
Секреция сока происходит непрерывно. Концентрированные корма и свекла способствуют наибольшему повышению секреции. В опытах «мнимого кормления» было установлено, что акт еды усиливает непрерывную секрецию сычуга. Усиление секреции наступает через 5—10 мин после начала кормления. Возбудителями секреции из изолированного желудочка являются 5%-ный раствор соды, 2%-ный раствор уксусной кислоты, пастой сена, 10—20%-ный раствор алкоголя при введении их в сычуг. Жир тормозит секрецию сычуга.
Вопрос о механизме возбуждения сычужных желез еще недостаточно изучен, однако и на основании полученных данных можно говорить о наличии у жвачных животных сложнорефлекторной и нервно-химической фаз секреции.
После механической и химической обработки пища из желудка поступает в кишечник, где идет дальнейший процесс пищеварения.
Важную роль в общем процессе пищеварения играет рубец. В нем сильно развиты бродильные процессы, вызываемые бактериями и симбиотическими инфузориями. По некоторым данным, количество последних достигает в содержимом рубца до 2000 в 1 см3.
Важное значение их в процессе пищеварения определяется тем, что они вызывают расщепление большого количества клетчатки, являющейся не только источником энергии, но и стимулятором моторики преджелудков. Кроме того, в результате переваривания самих инфузорий организм животного получает за сутки около 30—40% суточной потребности животного в белке. Процессы брожения в рубце ведут к образованию молочной, уксусной, пропионовой и масляной кислот, а также газов. При этом температура содержимого рубца повышается до 40—41° С. Белки перевариваются в незначительном количестве, главным образом в связи с действием ферментов бактерий, рН содержимого рубца колеблется в пределах 6,5—7,4. В рубце бактерии осуществляют синтез витаминов. Важную роль играют летучие жирные кислоты, образующиеся в рубце. Всасывание их происходит также в рубце, а затем они используются как основной источник энергии и для процесса образования молока. В результате интенсивных бродильных процессов в рубце накапливается большое количество газов, которые отрыгиваются по мере накопления. Отрыгивание газов — рефлекторный акт.
Желудочное пищеварение у птиц
У большинства птиц изо рта пища поступает в зоб, который является расширением пищевода. Зоб особенно развит у зерноядных птиц (куры, голуби). В зобе пищевая масса находится несколько часов, подвергаясь размягчению водой; слизистая зоба не содержит пищеварительных желез. Размягченная пища затем периодическими сокращениями мышц стенок зоба переводится в нижний отрезок пищевода, а оттуда в желудок. Последний имеет два отдела: железистый и мышечный (рис. 30). Первый содержит железы, выделяющие кислый желудочный сок с ферментами (пепсин, липаза).
Секреция желудочного сока у птиц идет медленно. Так, И. С. Цитович в опытах на курах, петухах и утках получал через фистулу отделение сока в количестве 0,8—3,8 мл за 5 мин. Кислотность сока колебалась от 0,003 до 0,4%; переваривающая сила равнялась 4 мм. Вид корма вызывал у петуха резко выраженную двигательную пищевую реакцию, повышение кислотности и переваривающей силы сока. При мнимом кормлении у гуся выделяется сок, содержащий 0,27% соляной кислоты. Однако имеется и химическая фаза. Содержимое желудка у гусей, уток и кур имеет кислую реакцию (рН = = 1,92—5,09).
Пребывание пищи в железистом отделе желудка птиц непродолжительно. Смоченная желудочным соком пища поступает в мышечный желудок, обладающий сильной мускулатурой и твердой слизистой оболочкой. Здесь происходит тщательное перетирание и переваривание желудочным соком пищи. Этому способствует наличие в желудке проглоченных камешков и песка. Сила мышечных сокращений желудка бывает настолько велика (у гусей — 265 мм рт. ст.), что по заключению некоторых исследователей, может раздробить стеклянные бусы и согнуть небольшие металлические пластинки (табл. 11).
Таблица 11. Внутрижелудочное давление у птиц при сокращении мускулатуры желудка (по X. С. Коштоянцу)
| ОбЪСКТ ИССЛСДОВаНИЯ | Давление В ЖСЛУДКС, ММ РТ. СТ |
| Гусь | 1 00—1 50 |
| Курица | 1 80 |
| Утка | 265—2 86 |
Возникновение сокращений мускулатуры желудка и пищевода связано с раздражением механорецепторов, заложенных в их стенках. Движения желудка и пищевода регулируются центральной нервной системой. Проводящими путями являются блуждающие и симпатические нервы. После механической и химической обработки пища эвакуируется из желудка в кишку, где происходит дальнейший процесс переваривания ее, в принципе не отличающийся от процессов переваривания у млекопитающих животных.
Желудочное пищеварение у свиней
С помощью фистульной методики, образования у свиней изолированного желудочка по И. П. Павлову и Гейденгайну, а также полизонда (по А. В. Квасницкому) изучены закономерности в работе желудочных желез, особенностью которых является их непрерывная деятельность. Кислотность желудочного сока колеблется в пределах 0,35— 0,45%. Прием корма усиливает сокоотделение в желудке. У свиней в желудке происходят бродильные процессы с образованием молочной кислоты. Возбудителями желудочной секреции являются корма растительного и животного происхождения. Механическое раздражение пустого желудка усиливает желудочную секрецию (Н. Ю. Беленков и В. Лосев). Регуляция деятельности желез желудка осуществляется так же, как и у собак, нейрогуморальным путем.
Желудочное пищеварение у лошадей. У лошадей с басовской фистулой и изолированным павловским желудочком в выделяемом желудочном соке определяли содержание соляной кислоты и ферментов. Исследованиями установлено, что кислотность составляет в среднем 0,24%, из которых 0,14% приходится на свободную соляную кислоту. В соке содержится фермент пепсин. Сокоотделение происходит в желудке лошади непрерывно и за сутки выделяется до 30 л сока. Прием корма усиливает секрецию сока. Сильные возбудители сокоотде ления: зеленый корм, клеверное сено, отруби, овес. Резко усиливает непрерывную секрецию желудочного сока механическое раздражение пустого желудка (С.С.Пол-тырев и др.). Регуляция желудочной секреции у лошадей в принципе аналогична той, которая происходит у собак. В растительном корме содержится большое количество клетчатки, которая не может перевариваться в желудке и тонком кишечнике, поэтому она подвергается перевариванию в слепой и большой ободочной кишках. Вместимость слепой кишки — 30—35 л. Так же, как и у жвачных, в слепой кишке сбраживание клетчатки происходит с участием микробов.
Глава 6
ПИЩЕВАРЕНИЕ В ТОНКОМ КИШЕЧНИКЕ
Из желудка пищевая масса поступает в двенадцатиперстную кишку, где подвергается химическому воздействию пищеварительных соков поджелудочной железы, печени и бруннеровского отдела кишечника. Получение дуоденального содержимого производится у животных через постоянную фистульную трубку, вставленную в кишку, а у человека с помощью тонкой резиновой трубки, вводимой через рот в двенадцатиперстную кишку.
Вне пищеварительного периода содержимое двенадцатиперстной кишки слабощелочной реакции: рН колеблется от 7,2 до 8,0. Секреция дуоденального сока производится бруннеровскими и либеркюновыми железами, расположенными в слизистой оболочке двенадцатиперстной кишки, напоминающими строение желез пилорической части желудка. Сок густой консистенции имеет щелочную реакцию. В нем содержится большое количество слизи, а также фермент (пептидаза), оказывающий переваривающее действие на белки. Некоторое действие дуоденальный сок оказывает на жиры и крахмал, кроме того, активирует фермент поджелудочной железы. Однако переваривающие свойства его сами по себе играют в пищеварительном процессе весьма ограниченную роль.
Верхний отдел двенадцатиперстной кишки играет важную роль в механизме регуляции секреторной и моторной деятельности пищеварительного аппарата, так как в его слизистой оболочке образуются гормоны: секретин, возбуждающий секрецию поджелудочного сока и желчи; холецистокинин, стимулирующий моторику желчного пузыря и угнетающий деятельность запирательного механизма общего желчного протока; вилликинин, возбуждающий моторику ворсинок тонкого кишечника; энтерогастрон, тормозящий секреторную функцию желудочных желез; «кишечное вещество», возбуждающее моторику кишечника, и др. Кроме того, этот отдел кишечника представляет собой мощное рецепторное поле, раздражение которого вызывает ряд физиологических реакций со стороны желудка, поджелудочной железы, печени и ее желчевыделительного аппарата, а также органов дыхательной, сердечно-сосудистой и кроветворной систем. Все это повышает значение дуоденального пищеварения. Оно усиливается еще и тем, что в полость этого отдела кишечника изливают сок, обладающий высокой пищеварительной активностью, секреторные клетки поджелудочной железы, а также желчь — клетки печени.
Внешнесекреторная функция поджелудочной железы
Поджелудочная железа (панкреас) — крупная пищеварительная железа, обладающая внешнесекреторной и внутрисекреторной функциями. Она является непарным органом и по строению напоминает слюнные железы. В поджелудочной железе различают головку, тело к хвост. Утолщенная правая часть поджелудочной железы расположена в петле двенадцатиперстной кишки, суженная левая часть — хвост — соприкасается с селезенкой. Тело железы имеет вид трехгранной призмы, спереди покрыто брюшиной.
Снаружи железа покрыта уплотненной соединительной тканью, от которой внутрь отходят перегородки, разделяющие ее на отдельные доли и дольки. Каждая долька имеет выводной проток, который впадает в более крупный междольчатый проток.
Междольчатые протоки, соединяясь, образуют главный выводной проток, открывающийся в двенадцатиперстную кишку. У некоторых животных и человека кроме главного панкреатического протока имеется добавочный проток, который самостоятельно впадает в двенадцатиперстную кишку.
Секреторные клетки железы имеют треугольную, цилиндрическую и округлую формы, в центре их расположено сферическое ядро. Кроме того, по всей железе рассеяны особые образования специальных клеток — островки Лангерганса, имеющие отношение к внутрисекреторной деятельности железы. Секрет этих клеток (инсулин) поступает непосредственно в кровь (Л. В. Соболев).
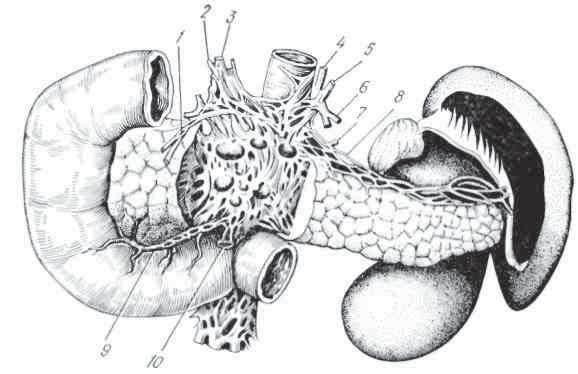
Рис. 31. Иннервация поджелудочной железы. Нервы проходят в сопровождении сосудов и только после отхождения внедряются в ткань железы (по Мюллер, 1924):
/ — артерия желудочпо-двекадцатиперстной кишки; 2 — правый чревный нерв: 3 —печеночная артерия; 4 — правый вагус; 5 — левый вагус; 5 —левая желудочная артерия; 7 — левый чревный нерв; 8— селезеночная артерия; 9 — внутренняя панкреато-дуоденальная артерия; 10 — верхняя брыжеечная артерия
Поджелудочная железа иннервируется блуждающими и симпатическими нервами и богато снабжена расположенными как в железистой ткани, так и в стенках протоков и сосудов интерорецепторами (рис. 31). Общая секреторная поверхность железы равна 11 м2; за час она способна выделять до 50 мл сока.
Методика исследования внешнесекреторной функции поджелудочной железы. Изучение внешнесекреторной функции поджелудочной железы у животных производится в остром и хроническом опытах. В остром опыте под общим наркозом производится лапаротомия и в главный проток поджелудочной железы вставляется стеклянная канюля, при помощи которой и собирают выделяющийся при различных условиях (раздражение нервов, вливание через специальную трубку различных веществ в двенадцатиперстную кишку) сок. По окончании опыта животное умерщвляется. В хроническом опыте животное подвергается операции: наложению хронической фистулы протока поджелудочной железы. Эта операция была в 1879 г. разработана И.П.Павловым. Она состоит в том, что из стенки двенадцатиперстной кишки вырезается участок с местом впадения главного панкреатического протока и вшивается в кожную рану (рис. 32). Дефект в кишке закрывается несколькими швами, и кишка подтягивается к кожной ране. Прооперированные таким образом животные живут в лабораторных условиях при специальном уходе долгие годы; выделяющийся у них секрет поджелудочной железы может быть в любое время подвергнут количественному и качественному анализу. Для сохранения нормального состояния животное получает весь оставшийся после анализа сок и, кроме того, к его пище добавляется небольшое количество соды.
А.Н.Бакурадзе предложил для получения поджелудочного сока у собак специальную металлическую канюлю, введенную в изолированный участок двенадцатиперстной кишки, куда открывается главный панкреатический проток. Проходимость кишки восстанавливается наложением кишечного анастомоза. Удобство этой операции заключается в том, что канюля дает возможность получать сок только во время опыта; вне опыта сок поступает через боковое отверстие фистульной канюли обратно в двенадцатиперстную кишку, благодаря чему исключаются хронические потери сока.
У людей получать чистый поджелудочный сок удавалось лишь В случаях образовавшегося наружного свища панкреатического протока, что бывает очень редко (рис. 33). На этих пациентах были проведены весьма важные физиологические исследования, которые легли в основу физиологии поджелудочной железы человека. Обычно же у человека исследование функции поджелудочной железы проводится при помощи тонкого зонда, вводимого через рот в двенадцатиперстную кишку — методика дуоденального зондирования. Однако получаемый при этом сок представляет собой смесь, состоящую из поджелудочного сока, желчи и соков бруннеровых и либеркюновых желез. К этой смеси может примешиваться и содержимое желудка, эвакуированное из желудка в двенадцатиперстную кишку
Рис. 32. Схема операции наложения хронической фистулы протока поджелудочной собаки (по И.П.Павлову)
Рис. 33. Больной с хронической фистулой протока поджелу- железы у дочной железы (по К.М. Быкову)
Для возбуждения секреции поджелудочного сока в двенадцатиперстную кишку обычно вводят 0,2—0,5% раствора соляной кислоты.
Состав и свойства поджелудочного сока
Чистый поджелудочный сок — бесцветная прозрачная жидкость щелочной реакции, без запаха, состоящая из неорганических и органических веществ. Из неорганических веществ большое значение имеет двууглекислый натрий, присутствие которого и обусловливает щелочность сока. Из органических — главную массу составляют белки. Содержание органических веществ колеблется от 0,5 до 8%; рН поджелудочного сока колеблется в пределах 8,71—8,98. Суточное количество выделяемого сока у собаки равно от 500 до 850 мл (по данным некоторых авторов 1000—1500 мл). У человека суточное количество сока достигает 600—850 мл (по данным некоторых авторов 1500—2000 мл).
В состав поджелудочного сока входят протеазы, липазы, амилаза, нуклеаза и другие ферменты. Амилаза, липаза, нуклеаза секретируются в активном состоянии, протеазы — в форме зимогенов, для перехода в активное состояние они нуждаются в воздействии других ферментов. Центральное место в процессе активации занимает трипсин, который активирует зимогены почти всех панкреатических ферментов — трипсиногена, химотрипсиногена, проэластазу и зимоген фосфолнпазы А. В отличие от всех других зимогенов, активация которых осуществляется трипсином, физиологическим активатором трипсиногена является фермент энтерокиназа, вырабатываемый слизистой оболочкой кишечника. Этот фермент был открыт Н. П. Шеповальниковым в лаборатории И.П.Павлова в 1899 г. Установлено, что местом наибольшей активности этого фермента является двенадцатиперстная кишка. Трипсин обладает наибольшей специфичностью и наибольшей скоростью гидролизовать аргинин, лизин, орнитин и др. Химотрипсин по сравнению с трипсином обладает более широкой субстратной специфичностью. С наибольшей скоростью он гидролизует связи, образованные карбоксильными группами. В поджелудочном соке в виде зимогена, активируемого трипсином, содержится эластаза.
Этот фермент обладает более широкой специфичностью по сравнению с трипсином и химотрипсином. В соке содержатся в виде зимогенов карбоксипептидазы А и В, они гидролизуют С-концевые аминокислотные остатки в молекулах белков и пептидов. В неактивном состоянии образуется в поджелудочной железе калликреин, при действии на глобулин плазмы он освобождает физиологически активный кинин. Активатором прокалликреина является трипсин, но он способен активироваться и спонтанно.
Амилаза поджелудочного сока сходна по своему действию с амилазой слюны. Рибонуклеаза расщепляет РНК до нуклеотидов.
Фосфолипаза А наибольшую активность проявляет по отношению к фосфолипидам анионного характера. Липаза гидролизует жиры.
Максимальное действие липазы на жиры проявляется при участии желчных кислот. Оптимум ее действия соответствует рН = 7,0—8,6.
Так же благоприятствует перевариванию жира находящийся в поджелудочном соке двууглекис лый натрий. Соляная кислота инактивирует липазу. Различные жиры расщепляются липазой с неодинаковой силой (табл. 12).
Таблица 12. Расщепление различных жиров под влиянием липазы поджелудочной железы
(по X.С. Коштоянцу)
| Жиры | Точка плавления‚°С | Растепление, % | Жиры | Точка плавления‚°С | Расщепление, % |
| Гусиный Куриный Бараний | 26—34 33—40 41-51 | 26,3 22,2 16,4 | Телячий Сливочное масло | 42—40 28-33 36-42 | 13,2 16,3 5,2 |
На различные пищевые вещества выделяется сок с различной концентрацией ферментов. Так, повышенная концентрация наблюдается в соке при еде молока и наинизшая — при еде мяса, причем отделение всех ферментов происходит параллельно. Наряду с этим наблюдения показали, что при приеме жирной пищи в желудочном соке увеличивается содержание липазы, при приеме углеводной — амилазы, белковой — трипсина.
Наиболее совершенным типом адаптации поджелудочной железы к режиму питания является, по-видимому, изменение количества ферментов. Менее экономной формой адаптации — увеличение количества секретируемого сока.
Отрицательное влияние на адаптационную способность поджелудочной железы оказывает резкий недостаток в рационе белка. Клиницистами отмечено, что у больных с заболеваниями желудочно-кишечного тракта способность поджелудочной железы к ферментным адап-тациям нарушается, что соответственно должно учитываться при назначении лечебного питания (А. Я. Губер-гриц, М. М. Губергриц).
Итак, физиологическое значение поджелудочного сока определяется: 1) участием в пищеварении, 2) нейтрализацией кислотности желудочного сока и 3) участием в регуляции кислотно-щелочного равновесия крови и водно-солевого обмена в организме.
Секреция поджелудочного сока. Секреторные клетки поджелудочной железы вне пищеварительного периода находятся в состоянии покоя и отделяют сок лишь в связи с периодической деятельностью желудочно-кишечного тракта. По данным лаборатории И.П.Павлова, у собак за каждый «период работы» выделяется в кишку около 30 мл смеси поджелудочного и кишечного соков с желчью. Периодическая секреция поджелудочного сока у человека, по данным П. И. Лепорского и его сотрудников, строго координирована с периодической деятельностью мускулатуры двенадцатиперстной кишки, и обычно моторика и секреция возникают одновременно. В период моторной деятельности кишки у здорового человека выделяется в среднем около 72,6 мл поджелудочного сока, богатого ферментами.
Во время приемов пищи происходит возбуждение секреторных клеток поджелудочной железы и наступает интенсивное и длительное выделение сока.
Наиболее обстоятельные и подробные данные о секреторной функции поджелудочной железы были получены И.П.Павловым и его сотрудниками. При приеме пищевых веществ было установлено следующее: 1) отделение поджелудочного сока происходит при виде и запахе пищи, т. е. в порядке условного рефлекса; 2) акт еды вызывает усиленное отделение сока, богатого ферментами; 3) латентный период сокоотделения на все сорта пищи колеблется от одной до трех минут; 4) работа поджелудочной железы типична для каждого сорта пищи; 5) концентрация основных ферментов (амилазы, липазы, трипсина) бывает наибольшей при еде молока и жира и наименьшей при еде мяса; 6) выделение всех ферментов идет параллельно; 7) концентрация ферментов в соке зависит от скорости секреции. Исследованиями была установлена зависимость количества отделяемого сока, хода и продолжительности секреции, а также количественного состава секрета от сорта пищи. При еде мяса или хлеба секреция характеризуется резким подъемом кривой в первые два часа, причем максимум отделения падает на второй час; в дальнейшем кривая отделения при еде мяса снижается и в четвертом часу резко падает; продолжительность отделения равна 4—5 ч. При еде хлеба снижение кривой начинается в третьем часу и затем секреция продолжается на низких цифрах еще около 5 ч; общая продолжительность отделения сока равна 9—10 ч. При приеме молока кривая секреции характеризуется максимальным подъемом в третьем часу, после чего происходит снижение; общая продолжительность секреции равна 5 ч (рис. 34).
По количеству выделяющегося сока при приеме различных сортов пищи на первом месте стоит хлеб, а на последнем — молоко.
Как показали наблюдения К. М. Быкова и его сотрудников на больном с хирургической фистулой протока поджелудочной железы, наибольшее количество сока у человека выделяется при приеме жирной пищи (табл. 13).
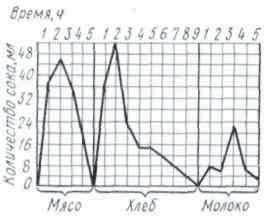
Рис. 34. Секреция поджелудочного сока у собаки при еде мяса, хлеба и молока (по И.П.Павлову)
Таблица 13. Секреция поджелудочного сока у человека при еде мяса, хлеба и молока (по К. М. Быкова)
Часы | Количествосока | ||
мясо (200 г) | хлеб (400 г) истакан чая | молоко(300мл) | |
1-й | 5,0 | 7,8 | 4,0 |
2-й | 5,2 | 5,3 | 4,0 |
3-й | 4,2 | 3,2 | 3,0 |
4-й | 2,1 | 3,4 | 4,5 |
При длительном приеме жирной пищи у человека происходит постепенное угнетение секреторной функции поджелудочной железы (табл. 14).
Таблица 14. Секреция поджелудочного сока у человека при прием в течение пяти дней жирной пищи (по К. М. Быкову)
Секреции за сутки | Количество сокав сутки, мл | ||||
первые | вторые | третьи | четвертые | пятые | |
День Ночь | 90,5 16,0 | 88,0 1,9 | 80,8 2,0 | 52,5 6,0 | 36,0 3,0 |
Всего | 106,5 | 89,9 | 82,8 | 58,5 | 39.0 |
Консистенция и объем пищи, а следовательно, и степень растяжения стенки желудка пищевыми массами играют большую роль в механизме возбуждения поджелудочной железы. Между количеством съеденной пищи и количеством отделяющегося при этом сока существует прямая зависимость: чем больше пищи поступает в желудок, тем сильнее секреция. Грубая пища, как, например, хлеб и картофель, возбуждает обильную секрецию поджелудочного сока.
Таблица 15. Содержание ферментов в поджелудочном соке собаки
Раздражители | Ферменты | |
амилаза, ед. | трипсин,мм | |
| Молоко | 256 | 6 |
| Хлеб | 256 | 4 |
| Мясо | 32 | 2,5 |
| Кислота | 64 | 2 |
| Механический | 256 | 5 |
Раздражение механорецепторов желудка в течение 3—4 ч вызывает через 1—2-минутный латентный период отделение поджелудочного сока. Секреция начинается интенсивно и в первый час достигает максимума, в дальнейшем происходит ее снижение, и к концу второго и началу третьего часа идет на сравнительно низком уровне. С прекращением раздражения секреция также прекращается. Повторное раздражение вновь вызывает интенсивное отделение сока. По качественному составу сок, полученный при раздражении механорецепторов желудка, больше соответствует соку, выделяющемуся на хлеб и молоко (табл.15).
Раздражение механорецепторов желудка возбуждает секреторную функцию поджелудочной железы у человека и одновременно повышает возбудимость ее на последующее действие пищевых раздражителей (табл. 16).
Во время секреции в железе резко повышаются окислительно-восстановительные процессы, о чем говорят увеличение потребления кислорода (в 2—3 раза) и повышение температуры железы. Сам секреторный процесс не является результатом простой фильтрации, а представляет собой активный процесс. И.П.Павлов, измеряя давление в кровеносном сосуде и протоке поджелудочной железы, установил, что при раздражении блуждающего нерва давление в сосуде было ниже, чем в протоке железы, однако секреция сока при этом была достаточно интенсивной. Сокоотделение, возникающее при раздражении блуждающего нерва, наблюдалось и в том случае, когда сосуды железы суживались под влиянием раздражения седалищного нерва.
Таблица 16. Влияние раздражения механорецепторов желудка на возбудимость поджелудочной железы к пищевым раздражителям
Условия опыта | Количествоподжелудочного сока, мл | |||
часы | ||||
1-й | 2-й | 3-й | всего | |
| Еда хлеба (250 г) Еде хлеба (250 г) предшествовало часовое механическое раздражение желудка | 60 77 | 55 63 | 33 33 | 148 173 |
При секреции сока в железе происходят морфологические изменения: количество секреторных зерен, которыми бывают насыщены клетки железы, становится значительно меньшим и они постепенно перемещаются к участку цитоплазмы, обращенному к просвету протока (табл. 17).
Таблица 17. Секреция поджелудочного сока у человека при раздражении механорецепторов желудка
| Секреция | Количество сока,мл за 1 ч | Ферменты | |
амилаза, ед. | трипсин, мм | ||
| «Периодическая» «Механическая» | 34 64 | 32 2048 | 2 11 |
Высокая внешняя температура вызывает уменьшение секреторной деятельности поджелудочной железы и снижение ферментативной активности сока не только в день нагревания, но и в течение нескольких дней (И.П.Разенков).
Возбудители поджелудочной секреции. Натуральными возбудителями поджелудочной железы, число которых сравнительно невелико, являются соляная кислота, овощные соки, клюквенный морс, жир и продукты его расщепления. Сильнейшим возбудителем является соляная кислота, менее энергичными — нейтральный жир, глицерин и высшие жирные кислоты. Вода относится к слабым возбудителям секреции поджелудочного сока. Энергичным возбудителем является механическое раздражение, оказываемое пищевой массой на рецепторный аппарат желудка. Из фармакологических веществ соко-гонным действием обладают алкоголь, эфир и пилокарпин. Среди перечисленных возбудителей секреции наибольший интерес представляет соляная кислота, которая является связующим звеном желудочного и поджелудочного пищеварения. Что касается пищевых возбудителей, то они приобретают особое значение при патологии желудка, в частности при ахилии, т. е. когда желудочный сок лишен свободной соляной кислоты. В этих случаях вода, жир и другие вещества целиком обусловливают работу поджелудочной железы.
По качественному составу сок, отделяющийся на кислоту, отличается от сока, отделяющегося на жир, воду и высшие жирные кислоты, тем что он беден органическими веществами и ферментами, но обладает высокой щелочностью с большим содержанием солей.
Механизм регуляции отделения поджелудочного сока. Механизм возбуждения поджелудочной железы является нервно-гуморальным и поэтому в ее деятельности так же, как и в деятельности желудка, различаются две фазы: сложнорефлекторная и нервно-химическая.
И.П.Павлов впервые установил, что блуждающие нервы, так же как и симпатические, содержат секреторные волокна для поджелудочной железы. В подтверждение этого им было представлено три доказательства: 1) увеличение секреции сока и повышение концентрации его основных компонентов при раздражении нервов, 2) паралич секреторных нервов при атропинизации и 3) независимость секреторного давления от давления крови.
И.П.Павловым было обнаружено в нервах, идущих к поджелудочной железе, наличие как секреторно-возбуждающих, так и секреторно-задерживающих волокон.
Перерезка блуждающего нерва, или полная денервация поджелудочной железы, вызывает резкие изменения ее секреторной функции, особенно со стороны количественного состава сока.
В лабораториях И.П.Павлова было установлено, что отделение поджелудочного сока при виде и запахе пищи, а также во время акта еды происходит рефлекторным путем. Точно так же и возбуждение поджелудочной железы при механическом раздражении желудка поступающей в него пищей осуществляется рефлекторным путем при участии блуждающих нервов и высших отделов центральной нервной системы. Кроме того, в этом случае передача возбуждения с желудка на поджелудочную железу может осуществляться и по «коротким путям», через местные интрамуральные механизмы.

В регуляции внешнесекреторной функции поджелудочной железы принимают участие подкорковые структуры, и в особенности гипоталамус (П.Г.Богач, В.Г.Томиленко, В. Губкин, П.С.Лященко). Так, раздражение передних и- средних участков гипоталамуса, включая серый бугор, вызывает выделение значительных количеств поджелудочного сока с большим содержанием плотного остатка за счет органических "веществ. Выделяющийся при раздражении передних и средних групп ядер гипоталамуса сок имеет высокую активность трипсина и амилазы. Раздражение задней части гипоталамуса вызывает слабое отделение сока. При раздражении гипоталамуса оральнее и на уровне воронки выделение сока наблюдается уже при действии раздражителя, а при раздражении серого бугра каудальнес воронки — в большинстве случаев после выключения раздражителя.
На секрецию оказывают влияние гормоны пищеварительного тракта — секретин, панкреозимин, гастрин, а также гормоны гипофиза, щитовидной железы, паращитовидных желез и надпочечников. Выяснено, что гипо-физэктомия снижает массу железы и секрецию бикарбоната и амилазы. Тироксин стимулирует синтез и секрецию амилазы, а кортизон—только синтез. Введение кортизона, АКТГ и альдостерона не оказывает влияния на секрецию, стимулированную субмаксимальными или максимальными дозами секретина у собак. Андреналэк-томия значительно снижает секрецию, а применение альдостерона частично ее восстанавливает. Антидиуретический гормон гипофиза снижает секрецию и концентрацию в соке бикарбоната при введении секретина.
Необходимо отметить, что вопросы влияния эндокринных желез на панкреатическую секрецию пока еще остаются мало изученными.
Наблюдениями за больными с фистулой протока поджелудочной железы, а также с неповрежденным желудочно-кишечным трактом, было установлено, что вид, запах пищи и разговор о еде вызывают у человека отделение сока, богатого ферментами (К. М. Быков и Г. М. Давыдов) (табл. 18). То же наблюдается при акте еды и во время раздражения механорецепторов желудка. При этих условиях возбуждение поджелудочной железы происходит рефлекторным путем, в порядке условных и безусловных рефлексов.
Исследованиями, проведенными в последнее время, было еще раз подтверждено условнорефлекторное возбуждение поджелудочной железы у человека. Так, здоровым и больным людям вводились в желудок и двенадцатиперстнуюкишку зонды, через которые периодически откачивалось содержимое. Было установлено: 1) акт еды вызывает секрецию и желудочного и поджелудочного сока; 2) по объему та и другая секреции превышают исходный объем на 30—50%; 3) содержание амилазы в дуоденальном содержимом при акте еды увеличивается в3 раза; 4) параллельно с увеличением секреции увеличивается концентрация соляной кислоты в желудочном соке и концентрация бикарбонатов в дуоденальном содержимом.
Нервно-химическая фаза поджелудочной секреции начинается с момента перехода кислого желудочного содержимого в двенадцатиперстную кишку. При действии соляной кислоты желудочного сока в слизистой оболочке двенадцатиперстной кишки происходит образование из просекретина гормона секретина, который через кровь поступает к поджелудочной железе и вызывает отделение поджелудочного сока.
Секретин — стойкое вещество, не разрушающееся при кипячении. В последнее время он был получен в кристаллическом виде, а также в виде хлористой соли. Секретин находится в слизистой оболочке кишечника человека, собаки, кошки, кролика и других животных (амфибий, рептилий, рыб, птиц). У беспозвоночных секретина, по-видимому, нет.
Доказательство «секретивного механизма» возбуждения поджелудочной железы было основано на следующих фактах: 1) введение соляной кислоты в денервиро-ванную двенадцатиперстную кишку вызывает секрецию поджелудочного сока; 2) при введении кислоты в двенадцатиперстную кишку возникает секреция сока из поджелудочной железы, пересаженной под кожу; 3) введение соляной кислоты в двенадцатиперстную кишку собаки, имеющей перекрестное кровообращение с другой собакой, вызывает секрецию поджелудочного сока не только у «донора», но и у «реципиента».
Раньше, на основании работ английских физиологов Бейлиса и Старлинга считали, что секретин действует непосредственно на секреторные клетки поджелудочной железы, без участия в этом процессе нервной системы. Однако последними экспериментальными исследованиями советских ученых было доказано, что в возбуждении поджелудочной секреции участвует симпатическая иннервация (А.В.Соловьев, X. С. Коштоянц, А.Н.Бакурадзе). Это получило подтверждение в опыте с введением в двенадцатиперстную кишку раствора соляной кислоты до и после подкожной инъекции эрготоксина — вещества, парализующего окончания чревных нервов (табл. 19). Из данных табл. 19 видно, что при предварительной инъекции эрготоксина введение в кишку соляной кислоты вызывает меньшую поджелудочную секрецию по сравнению с контролем. Однако приведенных данных недостаточно для утверждения, что секретин действует на поджелудочную железу через симпатическую нервную систему.
Таблица 19. Секреция поджелудочного сока на кислоту до и после введения под кожу эрготоксина (по А.В.Соловьеву)

Кроме гормона секретина в нервно-химической фазе секреции принимает участие и гормон панкреозимин-холецистокинин, также образующийся в слизистой оболочке двенадцатиперстной кишки. Этот гормон возбуждает ферментообразовательную функцию поджелудочной железы.
Помимо гормонов нервно-химическая фаза секреции обусловливается и некоторыми веществами, содержащимися в желудочном соке, а также продуктами переваривания пищи, которые после всасывания возбуждают через кровь нервно-железистый аппарат поджелудочной железы.
Сок, отделяющийся в период нервно-химической фазы, содержит меньшее количество органических веществ и ферментов и больше щелочей, чем сок, отделяющийся в период сложнорефлекторной фазы. Также различны и микроскопические изменения, происходящие в секреторных клетках при нервном и гуморальном раздражении поджелудочной железы.
В последние годы выяснено, что в клеточной мембране расположены рецепторы, с которыми взаимодействуют гормоны и медиаторы нервных импульсов. Все это является частью так называемой аденилциклазной системы. Вступая во взаимодействие с рецепторами, гормоны и медиаторы активируют аденилциклазу, которая затем активирует образование аденозинмонофосфата (АМФ) из АТФ. АМФ, в свою очередь, стимулирует внутриклеточные процессы и рассматривается как медиатор.
Итак, начальное возбуждение секреторных клеток поджелудочной железы происходит под влиянием услов-но-безусловнорефлекторных раздражений. Эфферентными проводниками импульсов являются блуждающие и чревные нервы. В дальнейшем сложнорефлекторная фаза сливается с нервно-химической.
Внешнесекреторная функция печени
Печень — крупная железа животного организма, участвующая в процессах пищеварения, обмена веществ, кровообращения и осуществляющая специфические защитные и обезвреживающие ферментативные и выдели тельные функции, направленные на поддержание постоянства внутренней среды организма. У взрослого человека ее масса достигает 1,5—2 кг. Печень вырабатывает пищеварительный секрет —желчь. Процесс образования желчи обозначается как секреция желчи, желчеобразование, или желчеотделение, а выход желчи в кишку — как желчевыделение. Желчеобразование и желчевыделение — тесно связанные процессы.
Строение печени и желчевыделительного аппарата. Печень —сложная трубчатая железа, состоит из двух долей, из которых правая значительно больше левой. Под серозной оболочкой располагается соединительнотканая капсула, содержащая эластичные волокна. Капсула вместе с кровеносными сосудами проникает внутрь печени, разделяя ее на печеночные дольки. В середине каждой дольки проходит центральная вена, от которой радиусом в виде перекладин располагаются печеночные клетки, между которыми образуются желчные капилляры. Таким образом, печеночные клетки находятся между разветвлениями кровеносных сосудов и желчными капиллярами (рис. 35).

Рис. 35. Строение печени. Вертикальный разрез печеночной балки (по Боор-геру):
I — печеночные клетки; 2 — желчные ходы; 3 и 5 — купферовы звездчатые клетки; 4 — лимфатическиепространства, 6 — кровеносные капилляры
Образовавшаяся в печеночных клетках желчь постепенно переходит к периферии дольки, где вначале поступает в междольчатые, а затем выводные печеночные протоки, которые вместе с пузырным протоком образуют общий желчный проток. У человека общий желчный проток открывается в полость двенадцатиперстной кишки обычно рядом с выводным протоком поджелудочной железы. На концевой части общего желчного протока находятся сфинктер Одди и интрамуральный запира-тельный механизм, содержащие циркулярные мышечные слои, и сфинктер ампулы, образующийся благодаря наличию циркулярной и продольной мускулатуры вокруг самой ампулы.
Вне периода пищеварения желчь из печеночных протоков через пузырный проток поступает в желчный пузырь; с началом пищеварения она через пузырный и общий желчный проток эвакуируется в кишку.
У человека желчный пузырь — тонкостенный грушевидный мешок—вмещает до 60 мл желчи; его длина равна 12—18 см; в нем различают дно, тело и шейку. Стенка его состоит из слизистой, мышечной и серозной оболочек. Постепенно сужаясь, желчный пузырь переходит в пузырный проток, который снабжен клапанами и на конце имеет сфинктер.
Методика исследования секреции желчи и выхода ее в кишку.
У животных изучение секреции желчи и выхода ее в кишку обычно проводится по методу хронического опыта. С этой целью делают постоянные фистулы желчного пузыря или общего желчного протока.
Желчеобразование изучается на животных с хронической фистулой желчного пузыря. При операции дно желчного пузыря подшивается к брюшной ране, после чего в его стенке делается надрез и слизистая пузыря выворачивается и подшивается к коже — получается грибовидный свищ желчного пузыря. Для получения всей желчи одновременно перевязывается общий желчный проток. Во избежание потери желчи вне опыта некоторые авторы предлагают вставлять в желчный пузырь металлическую фистульную трубку, как это делается при образовании фистулы желудка. Вне опыта фистула закрывается пробкой.
Выход желчи в кишку изучается при помощи выведения наружу устья общего желчного протока. И.П.Павлов разработал следующую операцию. У животного вырезается язычок из стенки кишки, куда впадает общий желчный проток, и пришивается к краям' кожной раны. Отверстие в кишке закрывается швами.
Г. В. Фольборт предложил операцию одновременного наложения хронической фистулы желчного пузыря и общего желчного протока по Павлову. При этих условиях у животного можно одновременно изучать желчеобразование и желчевыделение. С.М.Горшкова видоизменила операцию наложения общего желчного протока. Вместо вырезания кусочка кишки с протоком она иссекла цилиндрический

Рис. 36. Схема операции наложения хронической фистулы общего желчного протока и желчного пузыря у собаки (по С.М.Горшковой и Г. 3. Инасаридзе):
1 — печень; 2 — желчный пузырь; 3 — пузырный проток; 4 — печеночные протоки; 5 — общий желчный проток; 6 — малый панкреатический проток; 7 — изолированный отрезок двенадцатиперстной кишки; 8 — пилорическая часть желудка; 9 — большой панкреатический проток; 10 — двенадцатиперстная кишка; 11 — анастомоз; 12 — брюшная рана
отрезок кишки, один конец которого зашивался наглухо, а другой пришивался к краям кожной раны. Этим достигалось сохранение анатомии этой области и устранялось сдавлсние послеоперационными рубцами сфинктера Одди. Проходимость кишечника восстанавливалась при помощи гастроэнтероанастомоза, который позже был заменен энтеро-энтероанастомозом. Дополнительное наложение фистулы желчного пузыря позволяет так же, как и в методике Фоль-борта, одновременно изучать секрецию желчи и выход ее в кишку (рис. 36).
Для изучения желчевыдсления можно использовать канюлю Л. Н. Бакурадзе подобно тому, как это проводится при изучении функции поджелудочной железы, с той лишь разницей, что в данном случае следует иссекать цилиндрический отрезок двенадцатиперстной кишки с местом впадения общего желчного протока.
У человека исследование желчеобразовательной и желчевыдели-гельной функций печени обычно производится при помощи двух методов: дуоденального зондирования и холецистографии.
Метод дуоденального зондирования состоит в следующем. Из двенадцатиперстной кишки при помощи тонкого зонда извлекается кишечное содержимое золотисто-желтого цвета (желчь А); затем и кишку вводится 20—30 мл теплого 30%-ного раствора сернокислой магнезии или 30 мл 5—10%-ного раствора пептона, которые вызывают сокращение желчного пузыря и расслабление сфинктера Одди («желчный рефлекс»), благодаря чему густая темно-коричневая пузырная желчь (желчь В) поступает из желчного пузыря в кишку. При дальнейшем извлечении содержимое двенадцатиперстной кишки вновь становится светло-желтым (желчь С), эта желчь идет по печеночным протокам непосредственно из печени. Качественный анализ желчи А, В и С имеет большое клиническое значение, так как при этом определяется функциональное состояние печени и ее желчевыде-лительного аппарата.
В некоторых случаях получение желчи при дуоденальном зондировании производится путем подкожного или внутривенного введения пилокарпина, гипофизина или питуитрина. В последнее время предложен новый способ получения желчи у человека. Он состоит в том, что при дуоденальном зондировании производится фарадизация участка кожи — проекции прохождения правого диафрагмаль-ного нерва.
Метод холецистографии заключается в том, что внутривенно вводится раствор дериватов фенолфталеина (тетрабром или тетраиод гност), который, выделяясь печеночными клетками с желчью, обладает способностью задерживать рентгеновские лучи, это дает возможность определить при просвечивании контуры желчного пузыря, желчных протоков и их изменения. При обследовании больного всегда рекомендуется сделать холецистографию и дуоденальное зондирование.

Рис. 37. Графический метод регистрации сокращений желчного пузыря у человека и животных с хронической фистулой пузыря (по С.М.Горшковой):
1 — резиновый баллончик, вводимый через фистульное отверстие в полость желчного пузыря; 2 — резиновая трубка, соединяющая баллончик с градуированной бюреткой; 3 — зажим на трубке, через которую заполняются водой баллончик, бюретка и соединяющая трубка; 4 — градуированная бюретка, заполненная водой, по уровню которой определяется давление в пузыре (в мм вод. ст.); 5—поплавок, соединенный с писчиком 6. и производящий запись сокращений пузыря на кимографе 7; 8 — штатив
В физиологии рентгенографический метод применяется главным образом при обследовании моторной деятельности желчного пузыря, а отчасти с целью изучения желчевыделительной функции печени. В этих случаях перед рентгенографией в полость желчного пузыря вводятся различные контрастные вещества или к его стенкам прикрепляют металлические пластинки. Однако рентгенография дает представление лишь об изменениях формы и объема пузыря, обнаружить же свойственные пузырю перистальтические и ритмические сокращения при этом способе трудно.
Метод графической записи — более точный способ исследования. Он состоит в том, что животному через фистулу вводится в полость желчного пузыря небольшой резиновый баллончик, заполненный жидкостью и соединенный через водяной манометр с капсулой Марея, рычажок которой записывает на кимографе сокращения желчного пузыря. Вместо капсулы Марея С.М.Горшкова предложила записывать сокращения желчного пузыря при помощи специального рычажка, помещаемого на поплавок, находящийся в водяном манометре (рис. 37). Этот способ регистрации оказался очень удобным и более точно воспроизводящим сокращения мускулатуры пузыря. В клинике иногда удается проводить специальные физиологические исследования желчеобразовательной функции печени на людял, у которых свищи желчного пузыря, печеночных протоков и общего желчного протока образуются по медицинским показаниям операционным путем (камни желчного пузыря, опухоли общего желчного протока) или вследствие патологического процесса (воспаления).
Состав, свойства и физиологическое значение желчи.
Только что выделенная печенью желчь — печеночная желчь — жидкость светло-желтого цвета слабощелочной реакции. После пребывания в желчном пузыре, где она концентрируется, желчь становится густой, приобретает темно-коричневый цвет и содержит большое количество плотных веществ.
Основные составные части желчи: вода; желчные кислоты— гликохолевая и таурохолевая в форме натриевых солей; желчные пигменты — билирубин и биливердин; жиры и жироподобные вещества; холестерин, лецитин, слизь, хлористые, сернокислые и фосфорнокислые соли натрия, калия, кальция и магния. Из ферментов в желчи обнаружены фосфатазы.
Желчные кислоты и билирубин образуются в печени. Биливердин является продуктом окисления билирубина. Первый зеленого, а второй — красновато-желтого цвета. Билирубин образуется из гемоглобина при разрушении красных кровяных телец. Из желчных пигментов образуются пигменты мочи (уробилин) и кала (стеркобилин).
В настоящее время доказано, что все основные компоненты желчи секретируются. Полагают, что образование желчи начинается с активной секреции воды, желчных кислот и билирубина, осуществляемой паренхиматозными клетками печени. В результате в желчных канальцах появляется первичная желчь, которая, проходя по желчным ходам, вступает в контакт с плазмой крови, в результате чего устанавливается равновесие электролитов. Химическими исследованиями обнаружена наибольшая активность АТФ в мембранах желчных канальцев. Локализация кислой фосфатазы в печеночных клетках вокруг желчных канальцев указывает на функциональную полярность этих клеток в отношении желчной секреции. Энергия, необходимая для осуществления желчной секреции, образуется за счет тканевого дыхания печени и сопряженного с ним окислительного фосфори-лирования (А. С. Саратиков).
При некоторых болезненных состояниях организма билирубин, соединяясь с кальцием, образует нерастворимое вещество (билирубин-кальк), являющееся составной частью желчных камней (пигментные камни). В сутки у человека образуется от 800 до 1000 мл желчи.
Желчь принимает активное участие в процессе пищеварения. Она активирует ферменты поджелудочного и кишечного соков, главным образом липазу; эмульгирует жиры, ускоряя таким образом их переваривание; способствует растворению жирных кислот, обусловливая лучшее их всасывание; усиливает движение кишеч ника; возбуждает секрецию поджелудочной железы; тормозит развитие микробов, задерживая гнилостные процессы в кишечнике. Без желчи переваривание фибрина происходит лишь на 30,2%.

Рис. 38. Эвакуация желчи а кишку у собак при еде яичных желтков (А), молока (Б), мяса (Б), хлеба (Г) и раздражении механоре-цепторов желудка (Д). Числа — объем желчи в мл за 5 ч
Возбудители желчеобразования и желчевыделения. В настоящее время принято деление всех пищевых веществ на возбуждающие секреторную функцию печени (холеретика) и вызывающие выход желчи в кишку (хо-лагога). Однако такое разграничение является относительным, так как пищевые вещества не только вызывают выход желчи в кишку, но вместе с тем я являются энергичными возбудителями секреции желчи (рис. 38). Поэтому правильнее будет делить их на вещества с преобладающим действием на секреторный (желчесекре-торные вещества) и желчевыделительный аппараты печени (желчегонные вещества). Желчегонные вещества делят на усиливающие тонус желчепузырной системы — холекинетика (пептон, гистамин, холицнстокинин, яичный желток), и вещества, расслабляющие сфинктер Одди и интрамуральный механизм — холеспазмолитика (сернокислая магнезия, соляная кислота, питуитрин «Р», атропин). Некоторые из этих веществ, например пептон, холецистокинин, оказывают действие одновременно на концевой аппарат общего желчного протока и желчный пузырь. Энергичными возбудителями выхода желчи в кишку являются: яичный желток, жиры, пептоны, мясо, его экстрактивные вещества, хлеб, молоко, сыр; из химических веществ: пилокарпин, питуитрин, холецистокинин, ацетилхолин, физостигмин, гистамин, атофан, дюрандова смесь, сернокислая магнезия, бессмертник и каломель.
Слабыми возбудителями желчевыделения являются: сырой яичный белок, крахмал и алкоголь. Усиливают секрецию желчи соляная кислота, вода, желчь, секретин и механическое раздражение желудка. Ряд желчегонных веществ используются в клинике с диагностической целью (сернокислая магнезия, пептон, питуитрин и др.).
Секреция желчи и желчевыделение. Секреция желчи происходит непрерывно, выход же ее в двенадцатиперстную кишку связан с поступлением пищи в желудочно-кишечный капал. Исключением является периодическое желчевыделение во время голодания. При поступлении пищи в желудочно-кишечный капал начинается энергичная и продолжительная эвакуация желчи в двенадцати перстную кишку, причем в зависимости от сорта пищи (мясо, хлеб, молоко) желчевыделение характеризуется определенным латентным периодом, типичным ходом кривой и продолжительностью.
Латентный период выхода желчи в кишку у собак в среднем равен: на мясо — 5—8 мин, хлеб — 8—12 мин, молоко — 3—5 мин.
С.М.Горшковой в лаборатории К. М. Быкова было установлено, что величина латентного периода находится в обратной зависимости от степени наполнения желче-выделительного аппарата печени (табл. 20).
Таблица 20. Зависимость величины латентного периода желчевыделения от степени наполнения желчевыделительной системы печени (по С.М.Горшковой)
| Степень наполнения | Величина латентногопериода, мин | |||
молоко | мясо | хлеб | яичные желтки | |
Высокая Низкая | 3 23 | 8 40 | 12 55 | 5 25 |
Как показали наблюдения, желчный пузырь, который является главным «мотором эвакуации», начинает сокращаться в первые же минуты еды, это дает основание предполагать, что и у человека латентный период выхода желчи в кишку при еде различных видов пищи тоже незначителен. На основании холецистографии установлено, что поступление желчи в кишку у человека начинается
Через 5 мин после начала еды: через 10—15 мин объем желчного пузыря значительно уменьшается, а окончательное его опорожнение наступает спустя 3—5 ч.
У собак продолжительность желчевыделения различна и находится в зависимости от сорта пищи; так, при еде молока она равняется в среднем 63/4 ч, мяса – 51/2 ч, хлеба –81/4 ч.
Количество желчи, выделяющейся на эквивалентные по содержанию азота количества мяса, хлеба и молока, в среднем одинаково на все раздражители.
Установлено, что желчеобразовательная функция печени изменяется под влиянием различных пищевых режимов. Характер выхода желчи в кишку зависит от анатомических и физиологических особенностей организма (строение концевого аппарата общего желчного протока и желчного пузыря, возраст, беременность).
И.П.Павлов установил, что кривые желчевыделения и секреции поджелудочного сока при одинаковых сортах пищи в общих чертах сходны. Это свидетельствует об общности физиологической роли этих двух секретов в процессе пищеварения.
Желчевыделение происходит периодически, что обусловлено фазностью моторной деятельности желчевыде-лительного аппарата печени (П.К.Климов).
Роль желчного пузыря и общего желчного протока в эвакуации желчи в кишку. Главная роль в механизме эвакуации желчи в кишку принадлежит моторной деятельности желчного пузыря, сфинктера Одди и интраму-рального механизма общего желчного протока. Измерения внутрибрюшного давления, присасывающее действие двенадцатиперстной кишки, эластические свойства стенок желчного пузыря и секреторное давление печени, которым ранее приписывалась главенствующая роль, имеют лишь вспомогательное значение.
Вне периода пищеварения желчный пузырь человека находится в состоянии физиологического покоя и лишь иногда возникают единичные и слабые сокращения. У животных стенки желчного пузыря совершают непрерывные ритмические сокращения.
Способность желчного пузыря сокращаться была показана многими исследователями на различных животных (обезьяна, собака, кошка, кролик, морская свинка, птица, рыба, лягушка). Эта способность обусловлена наличием в его стенках гладких мышечных волокон и хорошо развитой системой нервных клеток. Кроме местных нервных ганглиев, обусловливающих автоматизм, движений желчного пузыря, в его стенках находятся окончания блуждающего и симпатического нервов, по которым к пузырю приходят импульсы из центральной нервной системы (рис. 39). Вид пищи и акт еды вызывают появление частых и сильных сокращений.
Сокращения желчного пузыря бывают трех типов: тонические, перистальтические .и ритмические. Частота и сила ритмических и перистальтических сокращений находятся в зависимости от качества возбудителя. Обычно частота ритмических сокращений в минуту достигает 5—7.
Возникновение топических сокращений также тесно связано с приемом пиши: вслед за быстрым 5—20-минутным подъемом тонуса наблюдается его резкое падение продолжительностью от 5 до 20 мин в зависимости от силы раздражителя; затем наступает новый подъём, продолжающийся 1—3 ч и более; при этом величина внутри-пузырного давления с 100—110 мм увеличиваете; до 240—370 мм вод. ст.

Рис. 39. Схема иннервации печени и желчных путей (по Мюллер, 1924):
1— серый бугор; 2 — ядро блуждающего нерва; 3 — левый вагус; 4 — правый вагус; 5 — верхний шейный узел; 6 — звездчатый узел, 7 — соединительные ветви; 8 — спинной мозг; 9 — большой чревный нерв; 10 — малый чревный нерв; 11 — чревный узел; 12 — печеночная артерия; 13 — печеночная ветвь вагуса; 14 — аорта; 15 — воротная вена; 16 — пищевод; 17 — пищеводный узел вагуса; 18 — желчный пузырь; 19 — печень; 20 — парасимпатические пути; 21 — симпатические пути
Максимальное внутрипузырное давление в норме обычно не превышает секреторного давления печени (табл. 21), чрезмерное повышение давления желчи в пузыре вызывает ослабление желчеобразования, вплоть до его полного прекращения. Регуляция давления по всей желчевыделительной системе осуществляется желчным пузырем, который обладает способностью концентрировать желчь в 10—20 и более раз. Моторным нервом желчного пузыря является блуждающий нерв. При раздражении симпатического нерва наблюдается торможение сокращений пузыря.
Таблица 21. Величина внутрипузырного и секреторного давления и человека и животных
| Объект исследования | Величинасопротивления кон-цевого аппаратаобщего желчного протока мм вод. ст. | Величинавнутрипузырного давления, мм вод.ст. | Величинасекреторного давления печени, мм вод. ст. |
Человек Обезьяна Собака Кошка Кролик | 100—230 100—300 100—300 30—75 30—150 | 100—370 100—350 100—350 100—125 70—100 | 200-385 230—321 110-342 150-260 100-125 |
В механизме регуляции моторной деятельности желчного пузыря участвует правый диафрагмальный нерв, раздражение которого усиливает сокращение стенок пузыря.
Деятельность желчного пузыря тесно связана с деятельностью сфинктера Одди, который так же, как и желчный пузырь, иннервируется от блуждающего, чревного и правого диафрагмалыюго нервов. Сравнительно недавно в концевой части общего желчного протока было установлено наличие второго сфинктера в интрамуральной части протока; сопротивление, создаваемое им току желчи, равно приблизительно 2/3 общего сопротивления в концевом аппарате общего желчного протока (300 мм вод. ст.).
У человека сопротивление сфинктеров в среднем не превышает 230 мм вод. ст. Однако в некоторых случаях сила их сокращения бывает настолько велика, что даже прием пищи не способен вызвать выхода желчи в кишку. При патологических процессах (дискинезии желчевыде-лительной системы) величина сопротивления значительно возрастает и может достигать 750—800 мм вод. ст.
На работу сфинктера не влияет тонус и перистальтика двенадцатиперстной кишки. Деятельность его изменяется при внутривенном введении пилокарпина, ацетилхо-лина и адреналина. Изолированный сфинктер способен реагировать на различные раздражители благодаря наличию в нем нервных сплетений.
Деятельность концевого аппарата общего желчного протока и желчного пузыря строго координирована, благодаря чему обеспечивается нормальное наполнение пузыря желчью и эвакуация ее в кишку. Наполнение желчного пузыря происходит при замыкании сфинктеров концевого отдела общего желчного протока и расслаблении мускулатуры стенок желчного пузыря, а эвакуация желчи в кишку происходит при расслаблении мускулатуры сфинктеров общего желчного протока и сокращении мышц желчного пузыря. Эта координация работы обусловлена регулирующим влиянием нервной системы.
В процессе наполнения пузыря и эвакуации желчи в кишку принимают участие мышечные элементы общего желчного протока, сфинктер и клапаны желчного пузыря.
Механизм регуляции секреции и выхода желчи в кишку. Процесс желчевыделения протекает в две фазы: сложнорефлекторную и нервно-химическую. В сложно-рефлекторной фазе принимают участие условные и безусловные рефлексы.
Первое указание на условнорефлекторное выделение желчи было сделано в лаборатории И.П.Павлова Г. Г. Брюно в 1898 г. Это положение в дальнейшем было подтверждено и значительно расширено.

Рис. 40. Условнорефлекторное возбуждение моторики желчного пузыря собаки. А — моторика пузыря, Б — дыхание, В — время: 1 — спонтанные сокращения после показа животному мяса; 2 — через 1 мин; 3 — через 3 мин; 4 — через 1 мин; 5 — через 3 мин; 6 — после 6 мин
Наиболее обстоятельные данные о положительном влиянии коры головного мозга на желчеобразовательную и желчевыделительную функции печени были получены советскими учеными. Было установлено, что секреция желчи и выход ее в кишку регулируются высшими отделами центральной нервной системы вид и запах так же, как и акт еды, способны усилить желчеобразование и вызвать выход желчи в кишку. Подобный эффект наблюдается при действии не только натуральных, но и искусственных условных раздражителей (рис. 40). Усиление секреции желчи и выход ее в кишку наступают рефлек-торно при поступлении пищевых и химических веществ в желудок и двенадцатиперстную кишку. Передача возбуждения с рецепторов желудка и кишки на печень происходит по системе блуждающих нервов и по «коротким путям» через местные интрамуральные механизмы. По симпатическим волокнам поступают тормозящие импульсы с рецепторов желудка к печени и желчевыделитель-ному аппарату, что при известных условиях может обусловить угнетение желчеобразования и прекращение эвакуации желчи в кишку.
Постоянство выхода желчи в кишку при введении в нее пищевых и химических веществ легло в основу клинических методов получения желчи у человека с диагностической целью («желчный рефлекс»).
В нервно-химической фазе желчевыделения принимают участие различные гормоны и продукты переваривания пищи.
Секретин стимулирует желчеобразовательную функцию печени, а выход желчи в кишку обусловлен действием специфического гормона — холецистокинина, который был найден в слизистой оболочке верхнего отдела тонкого кишечника у человека, а также различных животных (собаки, кошки, свиньи, овцы, коровы). Введение холецистокинина в организм вызывает сокращение желчного пузыря, расслабление сфинктера Одди и интраму-рального механизма общего желчного протока. Холецис-токинин — панкреозимин, как и все гормоны, действует на желчевыделительный аппарат при поступлении в кровь. При внутривенном введении человеку холецистокинина наблюдается эвакуация желчи в кишку. Подобный эффект отмечен и при инъекциях препаратов гипофиза.
В слизистой двенадцатиперстной кишки могут образовываться химические вещества, тормозящие желчеотделение. В механизме секреции желчи и желчевыделения принимают участие и сами продукты переваривания пищи, которые после всасывания оказывают действие на печень и желчевыделительный аппарат. Действие гормонов и химических веществ на желчевыделительный аппарат осуществляется при участии нервной системы.
В последние годы представление о механизме желчеобразования было значительно расширено в связи с учением о химической передаче нервного возбуждения. Так, при раздражении вегетативных нервов в крови, оттекающей от печени, были обнаружены медиаторы; введение холина и его производных или физостигмина вызывает усиление желчеобразования и выход желчи в кишку.
Значительное влияние на желчеобразование оказывают гормоны. Так, вазопрессин, АКТГ, инсулин его стимулируют, причем действие АКТГ осуществляется путем усиления продукции глюкокортикоидов. После гипофиз-эктомии, депанкреатизации или адреналэктомии желчеотделение уменьшается. То же происходит при введении препаратов щитовидной железы. Противоположный эффект вызывает тиреоидэктомия или длительное применение антитиреоидного препарата — 6-МТУ. Повышает уровень желчеотделения гормон эпифиза. После кастрации наступают длительные расстройства желчеотделения, что указывает на немаловажную роль половых гормонов в регуляции внешнесекреторной функции печени. Несмотря на разноречивые данные, можно полагать, что блуждающий нерв преимущественно усиливает желчеотделение, а симпатический тормозит.
На процесс желчеобразования оказывает влияние центральная нервная система. Еще в 1886 г. Гейденгайн установил, что раздражение электрическим током спинного мозга в шейном и верхнегрудном отделах вызывает снижение желчеобразования. То же наблюдается и при перерезке спинного мозга. Нервным центром регуляции желчеотделения считают центры блуждающих нервов. Имеются данные, что удаление мозговой коры у собак, повреждение премоторной и особенно лимбической области головного мозга изменяют желчеотделительную функцию печени (В. М. Бехтерев, Г. Е. Сабуров).
Все это дает основание говорить, что регуляция секреции желчи и выход ее в кишку осуществляются при помощи единого нервно-гуморального механизма, при ведущем значении нервного механизма.
Пищеварительная функция тонких кишок
В тонких кишках в основном завершается этап большой и сложной работы пищеварительного аппарата по механической и химической обработке поступившей е организм пищи.
В полость кишок изливаются поджелудочный сок, желчь и кишечный сок, обеспечивающие дальнейший гидролиз пищевых веществ при участии ряда ферментов. Гидролитические процессы завершаются здесь мембранным пищеварением. В тонкой кишке происходит всасывание почти всех продуктов гидролиза пищевых веществ, витаминов, большей части солей и воды.
Особое место в пищеварении и регуляции функций пищеварительного аппарата в комплексе с поджелудочной железой и печенью занимает двенадцатиперстная кишка, поскольку она является центральным узлом регуляции секреторной, моторной и эвакуаторной деятельности желудочно-кишечного тракта. Кроме того, в двенадцатиперстную кишку выделяются вещества гормональной природы, имеющие отношение к регуляции общего обмена веществ (Л. М. Уголен).

Рис. 41. Схема операции изолированной петли кишки по Тири — Велла
Тонкий кишечник у человека имеет в длину 6 м и более. На всем протяжении в слизистой оболочке его находятся либеркюновы железы, выделяющие за сутки до двух литров сока.
Исследование секреции кишечных желез обычно проводится в хроническом опыте на животных с постоянными кишечными фистулами. С этой целью животные подвергаются предварительной операции— изоляции отрезка кишки с сохранением сосудистой и нервной связи с организмом. Тири предложил иссекать небольшой отрезок тонкой кишки, один конец которой наглухо зашивается, а другой вшивается в кожную рану. Проходимость кишечника восстанавливается путем сшивания отрезков кишки «конец в конец» или «бок в бок». Велла несколько видоизменил эту операцию и стал оба конца изолированного отрезка кишки вшивать в кожную рану на небольшом расстоянии один от другого (операция Тири — Велла) (рис. 41). В других случаях ограничиваются просто вшиванием в кишку металлической трубки (канюли), как это делается при наложении басовской фистулы на желудок. Однако получить при этих условиях чистый секрет кишечных желез, конечно, невозможно. Такая фистула может служить лишь для анализа степени переваривания пищевой массы. Если такие фистулы наложить в различных отделах кишечника (полифистульный метод), то можно исследовать переваривание пищи по ходу кишечника (Е. С. Лондон).
В 1947 г. Л. Д. Синещековым была предложена методика, позволяющая направлять кишечный химус то в отверстие фистулы, то в нижележащие участки кишок. С помощью наружных анастомозов — мостиков можно во время опыта выключать большие участки кишечника.
Секреция бруннеровых желез
Бруннеровы железы расположены в начальном (проксимальном) участке двенадцатиперстной кишки. Сок их густой, сиропообразный, рН 7—8. Главным компонентом его является муцин, а пепсиноподобный фермент способен расщеплять белки, в кислой среде и медленно створаживать молоко. Секреция сока усиливается при приеме пищи и поступлении химуса из желудка.
Регуляция осуществляется нервно-гуморальным путем. Усиливает секрецию раздражение блуждающего нерва. Установлено участие холинергиче-ских нервных образований в регуляции сокоотделения. Стимулирующее влияние на секрецию оказывают секретин, глюкагон. Предполагают, что значение секрета бруннеровых желез защитное, так как густой сок, богатый муцином, может покрывать слизистую оболочку двенадцатиперстной кишки.
Кишечный сок и его свойства. Кишечный сок—мутноватая жидкость щелочной реакции, богатая ферментами и примесью слизи, эпителиальными клетками, кристаллами холестерина, микробами (небольшое количество) .и солями (0,2% углекислого натрия и 0,7% хлористого натрия). Железистым аппаратом тонкой кишки является вся его слизистая оболочка. Секреция кишечного сока включает два процесса: отделение жидкой и плотной частей сока. Плотная часть нерастворима в воде и представляет в основном отторгнутые от слизистой оболочки эпителиальные клетки. В ней содержится основная масса ферментов и других веществ (Г.К.Шлыгин). Жидкая часть — желтоватая жидкость щелочной реакции. В составе ее анионы и катионы, мукопротеин, мочевина, молочная кислота.
Содержание ферментов невелико. Кишечные ферменты, расщепляющие разные вещества, следующие: эрепсин — полип-ептиды и пептоны до аминокислот; катапепсины — белковые вещества в слабокислой среде (в дистальной части тонкого кишечника и толстых кишок, где под влиянием бактерий создается слабокислая среда); липаза — жиры на глицерин и высшие жирные кислоты; амилаза — полисахариды (кроме клетчатки) и декстрины до дисахаридов; маль-таза — мальтозу на две молекулы глюкозы; инвертаза — тростниковый сахар; нуклеаза — сложные белки (нуклеины); лактаза, действующая на молочный сахар и расщепляющая его на глюкозу и галактозу; щелочная фосфатаза, гидролизующая моноэфиры ортофосфорной кислоты в щелочной среде, кислая фосфатаза, обладающая тем же действием, но проявляющая свою активность в кислой среде, и др. Энтерокиназа, щелочная фосфатаза, выполнив свою роль в тонком кишечнике, толстых кишках, под влиянием микробов инактивируется (Г. К. Шлы-гин).
Содержание отдельных ферментов в кишечном соке варьирует в зависимости от различного питания. Так, например, при длительном нахождении животного па углеводном режиме питания содержание амилазы в кишечном соке увеличивается, а при длительном питании мясной пищей содержание ее уменьшается и возрастает количество ферментов, расщепляющих белки.
Секреция кишечного сока
Основными возбудителями секреции кишечного сока являются механические и химические раздражения, оказываемые пищевыми массами при прохождении через кишечную трубку. Механическое раздражение обусловливается самой массой, объемом пищи, имеющей различную консистенцию. Химическое раздражение производится желудочным и поджелудочным соками, желчью, а также продуктами переваривания белков, жиров и углеводов. При прохождении пищевой массы по кишечнику происходит раздражение механо-гермо-хеморецепторов стенок кишок; возникающие при этом импульсы рефлекторным путем передаются к кишечным железам, возбуждая их деятельность. В отличие от других пищеварительных желез кишечные железы отвечают на прием пищи не возбуждением, а торможением сокоотделения (В. В. Савич, М. П. Бресткин, И. М. Хазен и др.). Отсутствие усиления кишечной секреции па прием пищи пытаются объяснить тем, что слизистая оболочка тонкой кишки способна отделять сок по всей ее длине, а заполнение кишечника после приема пищи происходит постепенно. Если бы прием пищи сопровождался обильной секрецией, то во многих участках выделялось бы большое количество сока бесполезно в пустой просвет кишок. Это привело бы к неэкономной работе слизистой оболочки кишки. В связи с этим в процессе эволюции выработались механизмы регуляции, обеспечивающие выделение кишечного сока преимущественно при непосредственном контакте химуса со слизистой оболочкой.
Кишечные железы снижают секрецию не только во время приема пищи, но и при воздействиях на толстые кишки (П.Г.Богач, С.С.Полтырев).
Нервная система регулирует образование в железистых клетках ферментов. Она воздействует на железы не только во время секреции, но и в период относительного покоя железы, когда происходит интенсивное образование ферментов. Опыты с денервацией изолированной петли кишки показывают, что перерезка нервов влечет за собой увеличение количества сока с одновременным снижением в нем ферментов. Это указывает на нарушение трофического влияния на железистый аппарат.
В регуляции кишечной секреции участвуют и гормональные факторы. Стимулирующее влияние оказывают гормоны коры надпочечников. Оказывают влияние и половые гормоны: эстрадиол — пропинат стимулирует у самок, а тестеров— пропионат у самцов секрецию кишечного сока и особенно его плотной части. Введение половых гормонов усиливает преимущественно отделение плотной части сока и тем самым повышает его ферментативную активность.
Двигательная функция тонкого кишечника
Пищевая масса, поступая из желудка в кишечник, подвергается дальнейшей механической и химической обработке, в результате которой она приобретает гомогенный вид кашицы, называемой химусом.
Перемещение химуса по кишечнику происходит благодаря последовательным сокращениям заложенных в стенках кишечника продольных и кольцевых мышц.
Изучение двигательной функции кишечника производится при помощи следующих методик: 1) наблюдение за движениями кишок у животных со вскрытой брюшной полостью; 2) наблюдение за движением петли кишки, выведенной при помощи хирургической операции под кожу; 3) наблюдение за движением кишечника через вживленное в брюшную стенку целлулоидное окошечко; 4) графическая запись движений изолированного из организма и помещенного в питательный раствор Рингер — Локка или Тироде отрезка кишки; 5) графическая запись движений кишки при помощи резинового 'баллончика, введенного через фистулу в изолированную кишку по Тири, Тири — Велла или Павлову; 6) радиотелеметрическое исследование с помощью проглоченной радиокапсулы, имеющей датчик давления; 7) наблюдение за движениями кишечника при помощи рентгенографии. Последний метод широко применяется в клинике. При наличии у человека послеоперационных или травматических свищей кишечника изучение моторики последнего производится графическим методом.
В условиях хронических опытов чаще используется баллоногра-фическая методика, которая сравнительно проста и позволяет объективно регистрировать сокращения различных участков кишок (П.Г.Богач). Электрофизиологическими методами пользуются как в условиях хронических, так и острых опытов, а также in vitro на вырезанных отрезках кишки и полосках мышц. В хронических опытах регистрация биопотенциалов осуществляется при помощи элек-троэнтерографов или других электронных приборов с записывающим устройством. Отводящие электроды подшивают к мышечной оболочке со стороны серозного слоя или фиксируют их в подслизистом слое. В острых опытах и на отрезках кишки применяются различные электроды. Для внутриклеточного отведения потенциалов применяют стеклянные микроэлектроды.
Сокращения гладкой мускулатуры тонкой кишки неоднотипны, различают четыре типа: ритмическая сегментация, маятникообразные, перистальтические и антиперистальтические сокращения. Перистальтические бывают медленными и быстрыми (0,1—3,0 см/с). В проксимальных отделах тонкой кишки скорость их больше, чем средних, а в концевой части подвздошной кишки при стремительной перистальтике она достигает 7—21 см/с. Кроме перистальтики возникают тонические волны, продвигающиеся по кишке с меньшей скоростью. Могут возникать в кишке и непродвигающиеся тонические сокращения, которые обусловливают повышение тонуса кишки, сопровождаемое сужением просвета ее на довольно большом протяжении.
Перистальтические движения связаны с сокращениями кольцевой и продольной мускулатуры. Это длительные волнообразные сокращения и расслабления кишечника, напоминающие по форме движения червя, поэтому иногда их называют червеобразными. Происходят они таким образом, что кольцевая мускулатура довольно большого участка кишки сокращается при одновременном расслаблении мышц нижележащего участка. При этих движениях происходит проталкивание, выдавливание и передвижение химуса из верхних участков кишки в нижние. При некоторых состояниях организма, сопровождающихся рвотой, наблюдаются обратные перистальтические волны, при которых содержимое кишки направляется в сторону желудка (антиперистальтика). Ритми ческие движения связаны с сокращениями продольной и кольцевой мускулатуры сравнительно небольшого участка кишки (рис. 42). Они проявляются в том, что небольшой участок кишки то укорачивается, то удлиняется. По форме они напоминают движение часового маятника и поэтому их иногда называют маятникообразными движениями. Такая форма движений возникает в связи с тем, что происходит ритмическое сокращение и расслабление продольных мышц. Наряду с ритмическими сокращениями и расслаблениями продольных мышц происходят ритмические сокращения и расслабления кольцевых мышц этого же участка кишки, причем когда сокращается продольная мускулатура, то кольцевая расслабляется, и, наоборот, благодаря этому химус, находящийся в этом участке, то перемешивается, то передвигается в следующий участок. Эти своеобразные, строго координированные сокращения как бы расчленяют весь кишечник на отдельные небольшие сегменты, что дало основание физиологам называть их сегмента-ционными сокращениями.

Рис. 42. Моторика тонкой кишки у собаки
Ритм сокращений у собак в верхних отделах кишечника равен 18—20 цикл/мин, в нижних—12—14 цикл/мин.
На основании опытов с перерезками тонкой кишки было доказано, что на уровне впадения желчного и малого протоков поджелудочной железы локализуются датчики (водитель) ритма сокращений кишки, один из которых задает частоту сокращений верхним отделам тонкой кишки, а другой — в области подвздошной кишки.
Различные типы движений кишки в конечном счете обусловливают перемешивание химуса (что способствует перевариванию пищевых веществ и всасыванию продуктов их расщепления) и медленное передвижение его из верхних отделов тонкого кишечника к нижним отделам толстого кишечника.
Основными возбудителями движений кишки являются механические и химические раздражения, которые действуют на рецепторы, заложенные в стенках кишки.
Обычно чем грубее пищевой химус, тем энергичнее сокращается кишка. Сильными возбудителями моторики кишечника является растительная пища, содержащая в большом количестве клетчатку. Из химических раздражителей возбуждают сокращения кишки кислоты, введенные в кишечник в малых концентрациях, большие концентрации угнетают их. Оказывают действие щелочи, соли и другие вещества, особенно холин и его производные (ацетилхолин), образующиеся в кишечной стенке. По данным некоторых авторов, эти вещества считаются специфическими возбудителями моторики кишечника. Однако кишечник способен автоматически сокращаться независимо от влияний нервной системы. Поводом к такого рода заключению является то, что лишенная иннервации кишка совершает ритмическое движение. Этот автоматизм авторы связывают с функцией мускулатуры кишечника, признавая таким образом миогенное происхождение ритмической автоматики. В целостном же организме при нормальной иннервации кишечника все движения последнего подчинены регуляции со стороны цен-тральной нервной системы.
В регуляции двигательной деятельности тонкой кишки участвуют и интрамуральные нервные сплетения: ми-энтеральное (Ауэрбахово) глубокого межмышечного и подслизистого слоев (Мейсснера). В них происходит замыкание рефлекторных дуг.
Блуждающий и чревный нервы обеспечивают передачу тормозных и возбуждающих влияний на моторику кишки и афферентную сигнализацию из кишки в нервные центры. В составе блуждающего и чревных нервов содержатся возбуждающие и тормозные, а также афферентные волокна (В.Н.Черниговский, И. Л. Булыгин). Реакция на раздражение блуждающего нерва во многом зависит от исходного функционального состояния кишки в момент нанесения раздражения. Так, раздражение периферического конца перерезанного блуждающего нерва на фоне активных сокращений кишки вызывает торможение, а на фоне относительного покоя и слабых сокращений, наоборот, усиление моторики (Л. А. Коваль). Раздражение чревных и брыжеечных нервов чаще вызывает торможение всех видов сокращений кишок, реже возбуждение (Альварец).
В настоящее время П.Г.Богачем дана классификация моторных рефлексов кишечника.
1. Пищеводно-кишечный рефлекс, возникающий при повышении давления в полости пищевода или при раздражении его терморецепторов и сопровождающийся чаще всего повышением тонуса кишки. Передачу возбуждающего влияния осуществляется посредством блуждающих нервов, а тормозящего — чревных.
2. Желудочно-кишечный моторный рефлекс, возникающий под влиянием раздражения механорецепторов желудка и вызывающий усиление моторики кишки, причем передача осуществляется по стенке пищеварительной трубки, ее нервным сплетениям и рефлекторно посредством блуждающих нервов с замыканием рефлекторной дуги в центральной нервной системе. Имеются данные, указывающие на участие в передаче возбуждения с желудка на тонкую кишку чревных нервов.
3. Кишечно-кишечный моторный рефлекс, проявляющийся при действии механических и химических раздражений проксимального участка тонкой кишки усилением сокращения нижележащих отделов; при этом возбуждение передается как по стенке кишки в каудальном на-правлении, так и рефлекторно посредством блуждающих и чревных нервов.
4. Желудочно-цекальный рефлекс — один из разновидностей желудочно-кишечного моторного рефлекса, он обусловливает функциональную связь пилорического сфинктера и баугиниевой заслонки (И. И. Греков, И. М. Джаксон и др.).
5. Наблюдаемое после еды усиление сокращений конечной подвздошной (гастро-илеальный рефлекс) и толстой (желудочно-толстокишечный рефлекс) кишок — суммарный результат влияний с пищевода, желудка, двенадцатиперстной и тощей кишок, а также условно-рефлекторных влияний вида, запаха и вкуса пищи (П.Г.Богач, Е. А. Воробьев).
6. К тормозным рефлексам кишечника относится кишечно-кишечный (интестино-интестинальный, энтеро-эн-теральный) как частный случай кишечно-кишечного тормозного рефлекса. К тормозным рефлексам можно отнести ректо-энтеральный рефлекс (С.С.Полтырев с сотрудниками). Раздражение прямой кишки и анальных сфинктеров тормозит сокращения кишок. Замыкание дуги этого рефлекса происходит в спинном мозге (П.Г.Богач). Согласно данным П.Г.Богача, в осуществлении его участвуют волокна вагосимпатических стволов. В тормозном влиянии с прямой кишки определенную роль играет адреналин надпочечных желез.
С.С.Полтыревым был описан ректогастральный тормозной рефлекс. При сильном раздражении ампулы прямой кишки или анальных сфинктеров у собак наблюдается торможение желудочной секреции и моторики.
Согласно данным И.А.Булыгина с сотрудниками, висцеро-висцеральные рефлексы могут замыкаться вне центральной нервной системы: в узлах по ходу брыжеечных нервов, брюшных ганглиях и узлах симпатической цепочки.
П.Г.Богачем сформулирован основной закон рефлекторной регуляции моторики желудочно-кишечного тракта: адекватное раздражение любого участка желудочно-кишечного тракта вызывает возбуждение моторики в данном и нижележащих участках и усиление продвижения содержимого в каудальном направлении от места раздражения; одновременно оно тормозит моторику и задерживает продвижение химуса в вышележащих участках и отделах пищеварительного тракта.
Нервно-гуморальная регуляция моторики тонкого кишечника происходит при участии окситоцина, адреналина и норадреналина (катехоламины), оказывающих тормозящее действие. Ацетилхолин, в малых дозах серотонин и гистамин, гастрин I и II усиливают моторику, а питуитрин вызывает двухфазную реакцию — в начале кратковременное торможение, а затем следует усиление. Механизмы их действия пока изучены недостаточно, однако выяснено, что действие может осуществляться при участии интрамуральной нервной системы кишечника.

Рис. 43. Схема центральной регуляции моторной деятельности желудочно-кишечного тракта:
1 — передняя поясная извилина; 2 — орбитальная извилина; 3, 4 — височно-островковый участок; 5 — обонятельные доли; 6 — премоторная зона; 7 — грушевидная кора; 8 — гиппокамп; 9 — комплекс миндалевидных ядер; 10 — тала-мус и его ядра; // — передняя, средняя и задняя части гипоталамуса; 12 — средний мозг; 13 — гипофиз; /4 — ассоциативные связи; 15 — мозжечок; 16 — продолговатый мозг с ядрами блуждающих нервов; и — вагус; 18 — афферентные пути в грудном отделе спинного мозга от пищеварительного тракта и других органов; 19 — чревные нервы; 20 — надпочечник; 21 — брюшные ганглии (солнечное сплетение, верхний и нижний брыжеечные ганглии); 22 — влияние гипофизарных гормонов на кору надпочечников; 23 — влияние гормонов гипо-таламо-гипофизарного комплекса на пищеварительный тракт; 24 — желудок; 25 — тонкая кишка; 26 — адреналин, норадреналин, глюкокортикоиды и другие гормоны коры надпочечников; 27 — афферентные пути от внутренних органов; 28 — эфферентные пути (нервные и гуморальные) к дистальным отделам кишки и афферентные пути; анализаторы: 29 — обонятельный; 30 — зрительный; 31 — слуховой; 32 — вкусовой
В регуляции моторики участвуют высшие отделы центральной нервной системы — гипоталамус, структуры лимбической системы и кора мозга. В гипоталамусе локализованы центры, которые способны возбуждать или тормозить моторику. Благодаря нейросекреторной функции ядер гипоталамуса продуцируемые им гормональные вещества могут оказывать непосредственное действие на моторику. Согласно данным К. И. Несена, В.Д.Сокура, раздражение ядер передних и средних отделов гипоталамуса преимущественно возбуждает, а заднего — тормозит моторику желудка и кишок. Возбуждающий эффект наблюдается и при введении с помощью вживленных микроканюль непосредственно в структуру гипоталамуса адреналина и ацетилхолина. Необходимо отметить, что возникающие эффекты при воздействии на гипоталамус могут быть неодинаковыми в зависимости от локализации и силы раздражения, а также от исходного функционального состояния гладкой мускулатуры кишок. Кортикальные влияния осуществляются через лимби-ческую систему, но не исключено и непосредственное ее влияние на гипоталамус (рис. 43).
Экспериментальные данные свидетельствуют о том, что в коре содержатся механизмы, которые могут усиливать или ослаблять моторику. Корковый контроль за деятельностью пищеварительного тракта вполне возможен, если учесть наличие в коре представительств центров блуждающих и чревных нервов (В.Н.Черниговский). На важную роль коры мозга в регуляции моторики тонкой кишки указывают результаты клинических наблюдений К- М. Быкова с сотрудниками. Так, например, при отрицательном отношении пациента к лечебной процедуре у него регистрируется торможение моторики (А.В.Риккль и Е. П. Глинская). При действии натуральных положительных условных раздражителей наблюдается усиление моторики (рис. 44). Реакция кишки бывает неодинаковой в зависимости от вкусовых качеств принимаемой пищи. При неврозах нередко наблюдаются нарушения моторики кишок. Отрицательные эмоции (страх, испуг, беспокойство) сопровождаются торможением моторики. Во время сна моторика ослаблена.

Рис. 44. Усиление сокращений тонкой кишки человека при разговоре о бульоне (по К. М. Быкову и Г. М. Давыдову)
Периодическая деятельность пищеварительного аппарата
Сущность периодической деятельности пищеварительного аппарата (периодика) состоит в том, что при пустом желудке, отсутствии химуса в верхних отделах тонкой кишки или во всем тонком кишечнике у собак периодически возникает ряд секреторных и моторных явлений: сокращения желудка и кишечника, выделение поджелудочного и кишечного соков и желчи (период работы), которые затем сменяются периодами покоя. Эта периодика прекращается с началом еды и пищеварения. Патологические процессы и заболевания, в том числе и экспериментально воспроизведенные у собак, нарушают периодику (С.С.Полтырев с сотрудниками и др.).
При экспериментальной патологии коры головного мозга (неврозы) она также резко нарушается (И.Т.Курцин с сотрудниками).
Импульсы с коры мозга могут изменять чередование периодов покоя и работы (И. А. Булыгин).
Исследованиями установлено, что периодика свойственна человеку и ряду высших позвоночных животных.
Периодическая моторная деятельность желудка. До 1901—1902 гг. в лаборатории И.П.Павлова существовало мнение, что пищеварительные железы находятся обычно в полном покое и только под влиянием пищи, поступающей в ротовую полость начинается работа пищеварительного тракта.
Дальнейшие исследования привели к открытию периодической деятельности пищеварительного аппарата. Было установлено, что желудок вне пищеварительного периода совершает периодические сокращения, которые возникают через каждые 1,5—2 ч и продолжаются в течение 10—30 мин (рис. 45).
В дальнейшем было показано, что периодика появляется лишь на определенном этапе постнатального развития. У щенков она формируется в возрасте 1—2 месяцев, а периодика, свойственная взрослой собаке, устанавливается в разные сроки после рождения (желудка — в возрасте 6—9 месяцев, тонкой кишки — 10—16 месяцев).
У человека продолжительность периодов работы желудка 20—50 мин, покоя — 45—90 мин (С.В.Аничков, Карлсон, Н.И.Лепорский и др.).
Одновременно с сокращениями желудка наблюдаются сокращения нижней части пищевода и верхних отделов тонкой кишки (рис. 46). Периоды работы и покоя нижней части пищевода и желудка начинаются и заканчиваются почти одновременно (П.Г.Богач, Д.В.Филимонов и др.). Периодичность цикла, включающего период работы и период покоя, для желудка и верхних отделов тонкой кишки в основном одинакова по продолжительности. Период работы верхнего отдела тонкой кишки начинается одиночными сокращениями или небольшими группами с короткими паузами. Затем сокращения усиливаются и объединяются в более продолжительные группы, паузы между ними укорачиваются.

Рис. 45. Периодическая моторная деятельность желудка собаки вне пищеварения: А — Г — «периоды работы», чередующиеся с «периодами покоя»: Каждая последующая кривая — продолжение предыдущей
Согласно данным Н.Н.Лебедева, Т.Н.Клюбиной и др., периоды покоя и работы подвздошной кишки не совпадают с соответствующими периодами желудка и верхних отделов тонкой кишки. В периодике подвздошной кишки нет строгой закономерности, а часто она и вовсе отсутствует.
Моторная деятельность толстой кишки характеризуется у собак появлением серий сокращений различной длительности. Наряду с чередованием моторной активности и покоя можно наблюдать непрерывную моторику толстой кишки в течение 30—60 мин и более.
Периодика характеризуется не только двигательными, но и секреторными явлениями. В отсутствие кислой желудочной секреции периодически появляется выделение желчи и поджелудочного сока в кишку, секрета бруннеровых желез, кишечного сока и усиливается выделение слизи и сока пилорических желез. Компонентом периода работы пищеварительного аппарата у собаки является и слюноотделение. Особенностью секреторной деятельности поджелудочной железы у собак является то, что и в периоды покоя происходит выделение небольшого количества сока (Б.П.Бабкин, П.Г.Богач и др.). Выделяемый сок в периоды работы обладает высокой ферментативной активностью. Отделяемый кишечный сок содержит большое количество плотных веществ, ферментов высокой активности.
Во время периодов работы повышается тонус и усиливается сократительная и электрическая активность мышечной стенки желчного пузыря; соответственно повышается его внутрипузырное давление, что обеспечивает выход пузырной желчи в кишку, богатой холестерином, билирубином и холатами (С.М.Горшкова, И.Т.Курцин, А.В.Соловьев и др.). В периоды покоя эти показатели снижаются, однако может наблюдаться выделение небольшого количества желчи.
Важно отметить, что в норме у собак периодика при пустом желудке возникает, как правило, в отсутствии выделяемого кислого желудочного сока. Секреция кислого желудочного сока не является составным компонентом периодической работы пищеварительного аппарата.

При периодике пищеварительного аппарата возникают периодические изменения и в других функциональных системах организма. Так, например, в период работы желудка и тонкой кишки начинается тахикардия, усиливается кровоснабжение органов пищеварения, что является следствием повышения тонуса вазомоторного центра (Карлсон, Е.И.Синельников, В.Д.Суходоло). У человека повышается раздражительность, происходит усиление некоторых рефлексов. У животных отмечается активация поведенческих реакций, общедвигательное беспокойство, учащение дыхания и др. (И.П.Чукичев, А.И.Мордовцев, В.Д.Суходоло, К.В.Судаков и др.). Об изменениях в центральной нервной системе, возникающих одновременно с периодической активностью пищеварительного аппарата, свидетельствует происходящее изменение активности ядер блуждающих нервов (К.В.Судаков), появление периодических колебаний биоэлектрической активности рострального отдела ретикулярной формации и коры головного мозга. В крови наблюдаются периодические изменения содержания ацетилхолина, катехоламинов и холинэстеразной активности, причем в периоды работы содержание этих веществ и холинэстеразной активности повышается, а в периоды покоя — понижается (Е.Н.Сперанская, Е.М.Матросова). Все это свидетельствует о том, что периодическая деятельность не ограничивается только пищеварительным аппаратом.
Периодика органов пищеварения тесно связана с изменением обмена веществ в организме, причем изменения проявляются в периоды работы и покоя. Изменяются температура тела, легочная вентиляция, газообмен, основной обмен. В период работы эти показатели повышаются, покоя — снижаются. Во время периодов работы повышается активность многих ферментов, в том числе и окислительно-восстановительных, уровня сахара крови.
Таким образом, периодическая смена состояний работы состояниями покоя пищеварительного аппарата голодных животных тесно связана с периодическими изменениями обмена веществ и химического состава крови.
При кормлении животного (обычном или мнимом) во время периода работы сразу же наступает торможение движений желудка, которое затем сменяется перистальтическими сокращениями с появлением кислой желудочной секреции. Что же касается реакции тонкой кишки, то она протекает двухфазно: торможением сокращений и электрической активности со снижением тонуса кишки, которое затем сменяется усилением тонуса (П.Г.Богач, П.К.Климов и др.). Даже вид и запах пищи, поддразнивание собаки пищей мгновенно затормаживают сокращения пустого желудка (А.И.Мордовцев, А.Н.Бакурадзе, Н.Н.Лебедев) и верхних отделов тонкой кишки (П.Г.Богач).
Затормаживают периодику не только кормление собак, но и другие разнообразные факторы: введение в двенадцатиперстную кишку раствора соляной кислоты, адекватное растяжение стенок дистальных отделов тонкой кишки, илеоцекальной области, толстой и прямой кишок, механическое, химическое раздражение двенадцатиперстной кишки (С.С.Полтырев, А.В.Риккль, П.Г.Богач, И.П.Салмин, С.Д.Гройсман, Н.Н.Лебедев и др.). По выражению М.А.Собакина, каждый период работы пищеварительного аппарата у голодного животного — это пищеварительный период в миниатюре со всеми его функциями: секреторной (за исключением секреции кислого сока в желудке), моторной, всасывательной, эвакуаторной и экскреторной. Общий характер динамики его моторики сходен с динамикой моторики всего пищеварительного периода.
В согласованной деятельности различных частей пищеварительного тракта участвует центральная нервная система, местный нервный механизм и гуморальные факторы. Установлено, что перерезка вагосимпатических стволов на разных уровнях приводит к исчезновению периодики в желудке на длительный период, причем взамен ее возникают непрерывные сокращения редкого ритма. Меньше она отражается на двигательной периодике тонкой кишки, ибо приводит лишь к некоторому укорочению периодов работы и в течение нескольких первых недель к исчезновению или укорочению периодов покоя кишки. Существенные изменения претерпевает периодика после десимпатизации и при применении блокаторов симпатической иннервации (изобарипа, орнида). В частности, периоды покоя возникают в результате тормозных влияний из центральной нервной системы, передающихся по симпатическим нервам. Таким образом, в осуществлении периодики немаловажная роль принадлежит парасимпатической и симпатической иннервации.
Согласованная периодическая деятельность обеспечивается гипоталамусом как высшим центром регуляции вегетативных и некоторых соматических функций. Как указывает П.Г.Богач, во время физиологического голода гипоталамус действует как основной механизм деятельности разветвленной функциональной системы, направленной на выполнение поиска и потребления пищи, а также пищеварения. Воздействуя на гипоталамус, у животного существенно изменяется характер периодики. К центрам гипоталамуса при недостатке питательных веществ в нервные и другие ткани сигнализация поступает по нервным путям. Она, кроме того, может поступать от хеморецепторов сосудов и надгипоталамических структур мозга (П.Г.Богач, К.В.Судаков).
В настоящее время признано, что период активности пищеварительного аппарата обусловлен периодическими изменениями в деятельности гипоталамических механизмов, а эти изменения связаны с критическими изменениями содержания питательных веществ в крови и тканевых жидкостях.
Трансплантированный желудочек собаки проявляет периодику одновременно с большим (интактным) желудком (Фаррел и Айви). Это является свидетельством того, что в механизме регуляции периодики определенная роль принадлежит гуморальным факторам. Периодика сохранялась и в денервированном отрезке тонкой кишки. При введении катехоламинов в период работы происходит торможение сокращений желудка, а в период покоя — снижение тонуса его. Для протекания периодики определенное значение имеет серотонин, а также ряд гормонов. На периодике сказывается нарушение обмена веществ. Много исследований было посвящено выяснению физиологического значения периодики, высказывались различные мнения о се физиологическом смысле. Так, В.Н.Болдырев рассматривал периодику как процесс, необходимый для снабжения ферментами тканей организма со стороны поджелудочной железы. Это представление утратило свое значение. Б.П.Бабкин считал, что у голодного животного происходит накопление соков в протоках главных пищеварительных желез, а в период периодики они выводятся из протоков. По мнению И.П.Салмина, периоды работы возникают вследствие суммации возбуждений в пищевом центре, вызываемых импульсацией из пищеварительных желез при накоплении в них секретов на фоне тормозных влияний толстой кишки, когда возбуждение превалирует над тормозным влиянием со стороны толстой кишки, возникает разряд импульсов пищевого центра, которые по блуждающим нервам стимулируют деятельность пищеварительного тракта. Когда преобладают тормозные влияния с толстой кишки, наступает период покоя. По Салмину, главная роль -в механизме периодики принадлежит парасимпатическому отделу вегетативной нервной системы. Такое представление находится в несоответствии с современными данными. А.А.Лихачев и С.В.Аничков усматривали физиологическое значение периодики в экскреции накопившихся в крови ненужных или вредных продуктов для поддержания гомеостаза. Однако известно, что в составе соков содержатся также ферменты и другие ценные вещества. Кеннон и Карлсон связывали периодику с ощущением голода, считая, что сокращения желудка в периоды работы являются причиной ощущения голода, а последующая моторика и секреция — это подготовка пищеварительного тракта голодного животного для принятия и обработки пищи. Эта теория не получила должного подтверждения. Еще Шеррингтон в 1900 г. показал, что полная перерезка внешних нервов и удаление желудка не устраняют периодики. Можно лишь отметить, что действительно сокращения желудка в период работы могут усиливать чувство голода и импульсация из пищеварительного аппарата участвует в регуляции и формировании аппетита (В.Н.Черниговский, А.Н.Бакурадзе и др.). П.Г.Богач выдвинул теорию, по которой периодика у голодных животных и человека возникает в связи с переходом организма на эндогенный тип питания и рассматривается как необходимая и важная для сохранения нормальной деятельности организма. Первопричиной деятельности, направленной на удовлетворение потребностей организма в питательных веществах за счет собственных ресурсов, является состояние физиологического голода. Это состояние воспринимается гипоталамусом и другими структурами мозга, входящими в состав пищевого центра. Теория Богача, подкрепленная результатами многих экспериментов, получила довольно широкую известность и признание. Необходимо, однако, отметить, что поиск в выяснении физиологического значения периодической деятельности пищеварительного аппарата на этом не остановлен, он продолжается.
Глава 7
ПОЛОСТНОЕ И МЕМБРАННОЕ ПИЩЕВАРЕНИЕ
В тонком кишечнике существуют два взаимосвязанных типа пищеварения: полостное и мембранное (пристеночное). С помощью полостного пищеварения происходит первоначальный гидролиз пищевых веществ, на кишечной поверхности — его промежуточный и заключительный этапы.
Пристеночное пищеварение происходит на поверхности микроворсинок, расстояние между которыми колеблется примерно от 10 до 20 нм. В силу этого молекулы, размеры которых больше диаметра пор щеточной каймы, не могут проникнуть в последнюю, и мембранное пищеварение в отношении их будет неэффективно.
Учитывая, что животные и человек используют в качестве пищевых веществ ткани многоклеточных и одноклеточных организмов, то первоначальное полостное пищеварение является для них совершенно необходимым этапом переработки пищи. Пищевой материал, поступая из желудка в тонкую кишку, после частичного переваривания в желудке, недоступен действию ферментов, связанных со структурами кишечных клеток. На этом этапе более эффективны ферменты, действующие дистантно.
Таким образом, полостное пищеварение — наиболее эффективный механизм для гидролиза крупных пищевых молекул, клеточного материала. Мембранное пищеварение эффективно главным образом в отношении промежуточных продуктов гидролиза.
А. М. Уголев считает, что между полостным и мембранным пищеварением устанавливается положительная двусторонняя связь, обеспечивающая максимальную эффективность обоих механизмов. Полостное пищеварение подготавливает исходные субстраты для мембранного пищеварения; мембранное пищеварение уменьшает за счет перехода продуктов частичного гидролиза в щеточную кайму круг обрабатываемых субстратов
Пристеночное пищеварение осуществляется ферментами, фиксированными на клеточной мембране на границе внеклеточной и внутриклеточной сред. Пристеночное пищеварение характеризуется не близостью ферментативного гидролиза к поверхности клеточных мембран, а тем, что эти процессы происходят на мембранах, разделяющих две среды — внеклеточную и внутриклеточную, и тем, что его обусловливают ферменты, фиксированные на клеточных мембранах. Ферменты, действующие на поверхности тонкой кишки, имеют двоякое происхождение: частично они адсорбированы из химуса (амилаза, липаза, протеазы и т. д.), частично это собственно кишечные ферменты, синтезируемые внутри кишечных клеток и транслоцированные на поверхность клеточных мембран.
Продукты промежуточного гидролиза элиминируются в зону щеточной каймы, где происходит заключительная стадия гидролиза и переход к всасыванию. Расчеты показывают, что мембранное пищеварение осуществляет расщепление 80—90% пептидных и гликозидных связей, 55—60% триглицеридов.
В 50-х годах XX в. путем электронной микроскопии было установлено существование субмикроскопической
пористости, которая обусловливает увеличение активной поверхности и придает ей свойства пористого катализатора. То, что в световом микроскопе представлялось кутикулой, в действительности оказалось огромным количеством выростов клетки высотой около 0,62 мкм и диаметром около 0,08 мкм. Как выяснилось, на каждой клетке имеется около 3000 выростов (микроворсинок), а на 1 мм2 поверхности кишечного эпителия их около 50—200 млн. Расчеты показали, что благодаря наличию микроворсинок всасывающая поверхность кишки увеличивается в 14—30 раз. Микроворсинки — это цилиндрические выросты, ограниченные плазматической оболочкой, представляющие сплошные структуры цитоплазмы, покрытые снаружи типичной клеточной оболочкой толщиной около 10—12 нм (рис. 47). Высота их примерно в 7—10 раз больше диаметра.

Рис. 47. Схема кишечной клетки (по Триер, 1968):
1 — микроворсинки; 2 — терминальная сеть; 3 — десмосомы; 4 — митохондрии; 5 — гранулярный ретикулум; 6 — аппарат Гольджи; 7 — базальная мембрана; 8 — гладкий ретикулум; 9 — ядро
Размеры пор щеточной каймы приближаются к таковым многих пористых катализаторов, применяемых в промышленности. Значение катализатора состоит в том, что он увеличивает активную поверхность. Кроме того, они участвуют в переносе вещества в процессе работы катализатора в тех случаях, когда поры имеют приблизительно те же размеры, что и молекулы. Диаметр пор определяет предельную величину молекул, проникающих внутрь. Таким образом, микроворсинки превращают поверхность кишки в своеобразный пористый катализатор.
На основании электронно-микроскопических исследований предполагают, что микроворсинки способны сокращаться и расслабляться, что не может не влиять на протекание мембранного пищеварения. Отсюда следует, что размеры микроворсинок могут меняться в результате сокращения и расслабления.
В полости тонкой кишки осуществляются в основном начальные стадии, тогда как промежуточные и особенно заключительные происходят под влиянием ферментов, связанных со структурами эпителиальных клеток тонкой кишки Собственно кишечные ферменты в естественных условиях поступают в полость при десквамации кишечного эпителия. Время полного обновления кишечного эпителия у человека и млекопитающих колеблется в пределах от 6 до 14 дней. В общем балансе пищеварения действие ферментов, поступающих в полость тонкой кишки при десквамации кишечного эпителия, не может обусловливать значительный эффект. В процессах гидролиза пищевых субстратов участвуют не только слущенпые клетки, но и основная масса кишечных клеток в составе ворсинок.
В настоящее время имеются данные, свидетельствующие о том, что мембранное пищеварение регулируется. В частности, гормоны надпочечников, гипофиза и щитовидной железы оказывают стимулирующее влияние на синтез кишечных ферментов. Показано, что характер диеты оказывает значительное влияние на активность щелочной фосфатазы в щеточной кайме. Акт еды меняет ультраструктуру щеточной каймы и активность одного из локализованных в ней ферментов. Доказано, что состав ферментного слоя кишечной поверхности приспособлен к качеству питания.
Механизмы регуляции мембранного пищеварения связаны с моторикой желудочно-кишечного тракта (поскольку от нее зависит переход пищевых веществ из химуса в щеточную кайму); величиной пор щеточной каймы; интенсивностью потока жидкости внутрь кишечной клетки (должна отражаться на скорости поступления веществ в щеточную кайму); составом ферментного слоя; степенью поляризации мембраны и сорбционными свойствами мембраны.
Глава 8
ВСАСЫВАНИЕ В КИШЕЧНИКЕ
Всасывание — физиологический процесс, состоящий в том, что водные растворы питательных веществ, образовавшиеся в результате переваривания пищи, проникают через слизистую оболочку желудочно-кишечного канала в лимфатические и кровеносные сосуды. Благодаря этому процессу организм получает необходимые для жизни питательные вещества.
В верхних отделах пищеварительной трубки (рот, пищевод, желудок) всасывание весьма незначительное. В желудке, например, всасываются лишь вода, алкоголь, некоторые соли и продукты расщепления углеводов, причем в небольших количествах. Незначительное всасывание происходит и в двенадцатиперстной кишке.
Основная масса питательных веществ всасывается в тонком кишечнике, причем всасывание происходит в различных участках кишечника с неодинаковой скоростью. Максимум всасывания происходит в верхних участках тонких кишок (табл. 22).
Таблица 22. Всасывание веществ в различных отделах тонкого кишечника собаки
Всасываниевеществ в участке кишки, % | |||
Вещества | на25 см ниже | на100 смкверху | на2—3 см кверху |
привратника | отслепой кишки | отслепой кишки | |
Алкоголь | 70 | 18 | 0 |
Виноградныйсахар | 77 | 21 | 0 |
Крахмальный клейстер | 97 | 7 | 7 |
Пальмитиновая кислота | — | 37 | 12 |
Маслянаякислота | — | 16 | 2 |
В стенках тонкого кишечника имеются специальные органы всасывания — ворсинки (рис. 48).
Общая поверхность слизистой оболочки кишечника у человека равна приблизительно 0,65 м2, а вследствие наличия ворсинок (18—40 на 1 мм2) она доходит до 5 м2. Это приблизительно в 3 раза больше наружной поверхности тела. По Верцару, у собаки в тонком кишечнике имеется около 1 000 000 ворсинок.
Высота ворсинки 0,2—1 мм, ширина 0,1—0,2 мм, каждая содержит 1—3 мелких артерий и до 15—20 капилляров, находящихся под эпителиальными клетками. Во время всасывания капилляры расширяются, благодаря чему значительно увеличивается поверхность эпителия и его соприкосновение с протекающей в капиллярах кровью. В ворсинках имеется лимфатический сосуд с клапанами, открывающимися только в одном направлении. Благодаря наличию в ворсинке гладкой мускулатуры она может совершать ритмические движения, в результате которых происходит насасывание растворимых питательных веществ из полости кишки и выдавливание лимфы из ворсинки. За 1 мин все ворсинки могут всосать из кишечника 15—20 мл жидкости (Верцар). Лимфа из лимфатического сосуда ворсинки поступает в один из лимфатических узлов и далее — в грудной лимфатический проток.

Рис. 48. Поперечный срез тонкой кишки человека:
1 — ворсинка с нервным сплетением; г —центральный млечный сосуд ворсинки с гладкими мышечными клетка ми; 3 — либеркюновы крипты; 4 — mus-cularis mucosa; 5 — plexus submucosus; g _ submucosa; 7 — сплетение лимфатических сосудов; в — слой круговых мышечных волокон; 9 — сплетение лимфатических сосудов; 10 — ганглиозные клетки plexus myente; 11 — слой продольных мышечных волокон; 12 — серозная оболочка
После приема пищи ворсинки совершают движения в течение нескольких часов. Частота этих движений около 6 раз в 1 мин.
Сокращения ворсинок возникают под влиянием механических и химических раздражений, находящихся в полости кишечника веществ, например пептонов, альбумоз, лейцина, аланина, экстрактивных веществ, глюкозы, желчных кислот. Движение ворсинок возбуждается и гуморальным путем. Доказано, что в слизистой оболочке двенадцатиперстной кишки образуется специфический гормон вилликинин, который кровяным током подносится к ворсинкам и возбуждает их движения. Действие гормона и питательных веществ на мускулатуру ворсинок происходит, по-видимому, при участии нервных элементов, заложенных в самой ворсинке. По некоторым данным, в этом процессе принимает участие мейсснерогское сплетение, находящееся в подслизистом слое. При изоляции кишки из организма движения ворсинок прекращаются через 10—15 мин.
В толстом кишечнике всасывание питательных веществ при нормально-физиологических условиях возможно, но в незначительных размерах, а также веществ, легко расщепляющихся и хорошо всасывающихся. На этом основано в медицинской практике применение питательных клизм.
В толстом кишечнике довольно хорошо всасывается вода, в связи с чем кал приобретает плотную консистенцию. При нарушении в толстом кишечнике процесса всасывания появляется жидкий стул.
Е. С. Лондон разработал методику ангиостомии, при помощи которой удалось изучить некоторые важные стороны процесса всасывания. Эта методика состоит в том, что к стечкам крупных сосудов пришивается конец специальной канюли, другой конец выводится через кожную рану наружу. Животные с такими ангиостомическими трубочками живут при специальном уходе в течение долгого времени и экспериментатор, проколов длинной иглой стенку сосуда, может в любой момент пищеварения получить у животного кровь для биохимического анализа. Пользуясь этой методикой, Е. С. Лондон установил, что продукты расщепления белков всасываются по преимуществу в начальных отделах тонкого кишечника; всасывание же их в толстых кишках невелико. Обычно животный белок переваривается и всасывается от 95 до 99%, а растительный — от 75 до 80%. В кишечнике всасываются следующие продукты расщепления белка: аминокислоты, ди- и полипептиды, пептоны и альбумозы. Могут всасываться в небольшом количестве и нерасщеп-ленные белки: белки сыворотки крови, яичный и молока — казеин. Количество всасываемых нерасщепленных белков бывает значительным у детей раннего возраста (Р. О. Файтельберг). Процесс всасывания аминокислот в тонкой кишке находится под регулирующим влиянием нервной системы. Так, перерезка чревных нервов вызывает у собак усиление всасывания. Перерезка блуждающих нервов под диафрагмой сопровождается угнетением всасывания ряда веществ в изолированной петле тонкой кишки (Я- П. Скляров). Усиление всасывания наблюдается после экстирпации у собак узлов солнечного сплетения (Нгуен Тай Лыонг).
На скорость всасывания аминокислот оказывают влияние некоторые железы внутренней секреции. Введение животным тироксина, кортизона, питуитрина, АКТГ приводило к изменению скорости всасывания, однако характер изменения зависел от доз этих гормональных препаратов и длительности их применения (Н. Н. Калашникова). Изменяют скорость всасывания секретин и панкреозимин. Показано, что транспорт аминокислот осуществляется не только через апикальную мембрану энтероцита, но и через всю клетку. В этом процессе участвуют субклеточные органоиды (в частности, митохондрии). На скорость всасывания нерасщепленных белков влияют многие факторы и в частности патология кишечника, количество вводимых белков, внутрикишечное давление, избыточное поступление в кровь цельных белков. Все это может привести к сенсибилизации организма, развитию аллергических заболеваний.
Углеводы, всасываясь в виде моносахаридов (глюкозы, левулезы, галактозы) и отчасти дисахаридов, непосредственно поступают в кровь, с которой доставляются к печени, где они синтезируются в гликоген. Всасывание происходит очень медленно, причем скорость всасывания различных углеводов неодинакова. Если в стенке тонкой кишки моносахариды (глюкоза) соединяются с фосфорной кислотой (процесс фосфорилирования), всасывание ускоряется. Это доказывается тем, что при отравлении животного моноиодуксусной кислотой, тормозящей фос-форилирование углеводов, всасывание их значительно замедляется. Всасывание в различных участках кишечника неодинаково. По скорости всасывания изотонического раствора глюкозы отделы тонкой кишки у людей можно располагать в следующем порядке: двенадцатиперстная кишка > тощая кишка > подвздошная кишка. Лактоза в наибольшей степени всасывается в двенадцатиперстной кишке; мальтоза — в тощей; сахароза — в дистальной части тощей и подвздошной кишок. У собак участие разных отделов кишечника в основном такое же, как и у человека.
В регуляции процесса всасывания углеводов в тонкой кишке принимает участие кора головного мозга. Так, А. В. Риккль были выработаны условные рефлексы как на усиление всасывания, так и на задержку. Изменяется интенсивность всасывания при пищевом возбуждении, при акте еды. В условиях эксперимента удавалось влиять на всасывание углеводов в тонкой кишке путем изменения функционального состояния центральной нервной системы, фармакологическими средствами, раздражением током разных корковых областей у собак с вживленными электродами в лобную область, теменную, височную, затылочную и заднюю лимбнческую области коры головного мозга (Р. О. Файтельберг). Эффект зависел от характера сдвига в функциональном состоянии коры головного мозга, в опытах с применением фармакопре-паратов, от участков коры, подвергавшихся раздражению током, а также и от силы раздражения. В частности, выявлено большее значение в регуляции всасывательной функции тонкого кишечника лимбической коры.
Каков механизм включения коры мозга в регуляцию всасывания? В настоящее время имеются основания предполагать, что информацию в центральную нервную систему о происходящем процессе всасывания в кишечнике несут импульсы, возникающие как в рецепторах пищеварительного тракта, так и кровеносных сосудов, причем последние раздражаются поступившими из кишечника в кровяное русло химическими веществами.
Немаловажное участие принимают подкорковые структуры в регуляции всасывания в тонкой кишке. При раздражениях латеральных и задне-вентральных ядер зрительного бугра изменения всасывания сахара были неодинаковыми: при раздражении первых наблюдалось ослабление, при раздражении вторых усиление. Изменения интенсивности всасывания наблюдались при раздражениях бледного шара, миндалевидного тела и при раздражении током подбугровой области (П. Г. Богач).

Рис. 49. Нейроэндокринная регуляция процессов всасывания в кишечнике (по Р. О. Файтельбергу и Нгуен Тай Лыонгу): Черные стрелки — афферентная информация, белые — эфферентная передача импульсов, заштрихованные — гормональная регуляция
Таким образом, участие подкорковых образований в регуляции всасывания сахара можно считать доказанным. На всасывательную деятельность тонкой кишки оказывает влияние ретикулярная формация ствола мозга. Об этом свидетельствуют результаты опытов с применением аминазина, блокирующего адренореактивные структуры ретикулярной формации. В регуляции всасывания участвует мозжечок, способствующий оптимальному течению процесса всасывания в зависимости от потребностей организма в питательных веществах.
Согласно последним данным импульсы, возникающие в коре головного мозга и нижележащих отделах центральной нервной системы, достигают всасывательного аппарата тонкой кишки через вегетативный отдел нервной системы. Об этом свидетельствует тот факт, что выключение или раздражение блуждающих или чревных нервов существенно, но не однонаправленно изменяют интенсивность всасывания (в частности, глюкозы).
В регуляции всасывания участвуют и железы внутренней секреции. Нарушение деятельности надпочечников отражается на всасывании углеводов в тонкой кишке. Введение в организм животных кортина, преднизолона изменяет интенсивность всасывания. Удаление гипофиза сопровождается ослаблением всасывания глюкозы. Введение животному АКТГ стимулирует всасывание; удаление щитовидной железы снижает интенсивность всасывания глюкозы. Снижение всасывания глюкозы отмечается и при введении антитиреоидных веществ (6-МТУ). Имеются некоторые основания признать, что гормоны поджелудочной железы способны оказывать влияние на функцию всасывательного аппарата тонкой кишки (рис.49).
Нейтральные жиры всасываются в кишечнике после расщепления на глицерин и высшие жирные кислоты. Всасывание жирных кислот обычно происходит при соединении их с желчными кислотами. Последние, попадая в печень через воротную вену, выделяются печеночными клетками с желчью и таким образом могут опять принимать участие в процессе всасывания жиров. Всасываемые продукты расщепления жира в эпителии слизистой оболочки кишечника вновь синтезируются в жир.
Р. О. Файтельберг считает, что процесс всасывания состоит из четырех этапов: транспорта продуктов полостного и пристеночного липолиза через апикальную мембрану; транспорта жировых частиц по мембранам канальцев цитоплазматической сети и вакуоли пластинчатого комплекса; транспорта хиломикронов через боковые и. базальные мембраны; транспорта хиломикронов через мембрану эндотелия лимфатических и кровеносных сосудов. Скорость всасывания жиров зависит, вероятно, от синхронности работы всех этапов конвейера (рис. 50).
Установлено, что одни жиры могут влиять на всасывание других, а всасывание смеси из двух жиров происходит лучше, чем каждого в отдельности.
Всосавшиеся в кишечнике нейтральные жиры попадают в кровь через лимфатические сосуды в большой грудной проток. Такие жиры, как масло и свиной жир, всасываются до 98%, а стеарин и спермацет — до 9— 15%. Если у животного через 3—4 ч после приема жирной пищи (молока) вскрыть брюшную полость, то легко можно увидеть невооруженным глазом наполненные большим количеством лимфы лимфатические сосуды брыжейки кишечника. Лимфа имеет молочный вид и получила название млечного сока или хилуса. Однако не весь жир после всасывания поступает в лимфатические сосуды, часть его может направляться в кровь. В этом можно убедиться, если у животного перевязать грудной лимфатический проток. Тогда содержание жира в крови резко увеличивается.
Вода поступает в желудочно-кишечный тракт в большом количестве. У взрослого человека суточное потребление воды достигает 2 л. В течение суток у человека в желудок и кишечник выделяется до 5—6 л пищеварительных соков (слюны — 1 л, желудочного сока — 1,5— 2 л, желчи — 0,75—1 л, поджелудочного сока — 0,7— 0,8 л, кишечного сока — 2 л). Выводится из кишечника наружу только лишь около 150 мл. Всасывание воды происходит частично в желудке, интенсивнее в тонком и особенно толстом кишечнике.
Растворы солей, главным образом поваренной соли, всасываются довольно быстро, если они гипотоничны. При концентрации поваренной соли до 1% всасывание идет интенсивно, а до 1,5% всасывание соли прекращается.
Растворы солей кальция всасываются медленно и в незначительном количестве. При высокой концентрации солей происходит выделение воды из крови в кишечник. На этом принципе в клинике построено применение некоторых концентрированных солей в качестве слабительных веществ.
Роль печени в процессе всасывания. Известно, что кровь из сосудов стенок желудка и кишечника поступает через воротную вену в печень, а затем уже через печеночные вены в нижнюю полую вену и далее в общий круг кровообращения. Ядовитые вещества, образующиеся в кишечнике при гниении пищи (индол, скатол, тира-мин и др.) и всасывающиеся в кровь, обезвреживаются в печени путем присоединения к ним серной и глюкуро-новой кислот и образования мало ядовитых эфирно-серных кислот. В этом состоит барьерная функция печени. Выяснена она была И. П. Павловым и В. Н. Экком, которые на животных проделали следующую оригинальную операцию, получившую название операции Павлова— Экка. Воротная вена путем анастомоза соединяется с нижней полой веной, и таким образом кровь, оттекающая из кишечника, попадает в общий круг кровообращения, минуя печень. Животные после такой операции погибают через несколько дней вследствие отравления ядовитыми веществами, всосавшимися в кишечнике. Особенно быстро приводит животных к гибели кормление мясом.

Рис. 50. Механизм переваривания и всасывания жиров. Четырехэтапный транспорт липидов с длинными цепями через энтероциты (по Р. О. Файтельбергу и Нгуен Тай Лыонгу)
Печень является органом, в котором происходит ряд синтетических процессов: синтез мочевины и молочной кислоты, синтез гликогена из моно- и дисахаридов и др. Синтетическая функция печени лежит в основе антитоксической функции ее. При введении в желудочно-кишечный канал бензойнокислого натрия в печени происходит нейтрализация его путем образования гиппуровой кислоты, выделяемой затем из организма почками. На этом основана одна из функциональных проб, применяемых в клинике при определении синтетической функции печени у человека.
Механизмы всасывания
Процесс всасывания состоит е том, что питательные вещества проникают через клетки эпителия кишки в кровь и лимфу. При этом одна часть питательных веществ проходит через эпителий не изменяясь, другая — подвергается синтезу. Движение веществ идет в одном направлении: от полости кишки к лимфатическим и кровеносным сосудам. Это связано со структурными особенностями слизистой оболочки стенки кишки и составом веществ, содержащихся в клетках. Определенное значение имеет давление в полости кишечника, Которое отчасти обусловливает процесс фильтрации воды и растворенных веществ в клетки эпителия. При увеличении давления в полости кишки в 2—3 раза всасывание, например раствора поваренной соли, увеличивается в 2 раза.
В свое время считалось, что процесс фильтрации полностью обусловливает всасывание веществ из полости кишки в клетки эпителия. Однако такая точка зрения является механистической, поскольку рассматривает процесс всасывания, являющийся сложнейшим физиологическим процессом, во-первых, с чисто физических принципов, во-вторых, без учета биологической специализации органов всасывания и, наконец, в-третьих, в отрыве от всего организма в целом и регулирующей роли центральной нервной системы и ее высшего отдела — коры больших полушарий головного мозга. Несостоятельность фильтрационной теории видна уже из тех фактов, что величина давления в кишке приблизительно равна 5 мм рт. ст., а величина давления крови внутри капилляров ворсинок доходит до 30—40 мм рт. ст., т. е. в 6— 8 раз больше, чем в кишке. Об этом свидетельствует и тот факт, что проникновение питательных веществ при нормальных физиологических условиях идет лишь в одном направлении: от полости кишки к сосудам лимфы и крови; наконец, опытами на животных доказана зависимость процесса всасывания от кортикальной регуляции. Установлено, что импульсы, возникающие при условнореф-лекторном раздражении, могут то ускорять, то замедлять скорость всасывания веществ в кишечнике.
Несостоятельными и метафизическими являются и теории, объясняющие процесс всасывания только законами диффузии и осмоса. В физиологии накопилось достаточное количество фактов, противоречащих этому. Так, например, если ввести в кишку собаки раствор виноградного сахара в концентрации меньшей, чем содержание сахара в крови, то вначале происходит всасывание не сахара, а воды. Всасывание сахара в данном случае начинается лишь тогда, когда концентрация его в крови и полости кишки будет одинакова. При введении в кишку раствора глюкозы в концентрации, превышающей концентрацию глюкозы в крови, происходит вначале всасывание глюкозы, а затем уже воды. Точно так же, если ввести в кишку сильно концентрированные растворы солей, то вначале происходит поступление в полость кишки из крови воды, а затем, при выравнивании концентрации солей в полости кишки и в крови (изотония), происходит уже всасывание раствора солей. Наконец, если в перевязанный участок кишки ввести сыворотку крови, осмотическое давление которой соответствует осмотическому давлению крови, то вскоре же сыворотка полностью всасывается в кровь.
Все эти примеры свидетельствуют о наличии в слизистой оболочке стенки кишечника одностороннего проведения и специфичности для проницаемости питательных веществ. Поэтому объяснить явление всасывания исключительно процессами диффузии и осмоса нельзя. Однако эти процессы, несомненно, играют определенную роль при всасывании питательных веществ в кишечнике. Процессы диффузии и осмоса, протекающие в живом организме, коренным образом отличны от этих процессов, наблюдаемых в искусственно созданных условиях. Слизистую оболочку кишки нельзя рассматривать, как это делали некоторые исследователи, только лишь как полупроницаемую перепонку, мембрану.
Слизистая оболочка кишки, ее ворсинчатый аппарат представляют собой такое анатомическое образование, которое специализировано к процессу всасывания и функции его строго подчинены общим закономерностям живой ткани целостного организма, где любой процесс регулируется нервной и эндокринной системами.
Глава 9
ПИЩЕВАРЕНИЕ В ТОЛСТОМ КИШЕЧНИКЕ
На пути прохождения химуса из тонкого кишечника, в толстый имеется так называемая баугиниева заслонка, которая обладает высокой чувствительностью. При участии рефлекторного механизма она регулирует переход химуса из тонкого кишечника в толстый. Заслонка имеет мышцы, расположенные в различных направлениях, что обусловливает разнообразные виды ее движений. В течение 24 ч через заслонку проходит приблизительно от 2000 до 4000 г химуса. По данным Е. С. Лондона, у собак в толстую кишку переходит 10% принятых с пищей азотистых веществ, 5% крахмала и 3% жира.
Отделение сока в толстой кишке происходит непрерывно, хотя и в небольших количествах. Возбудителями его отделения являются механические и химические раздражения. Сок состоит из плотной части и жидкой. Плотная часть представляет собой серовато-желтоватые комочки с примесью слизи, содержит отторгнутые распадающиеся эпителиальные клетки плюс лимфоидные элементы. В плотной части сока содержатся в небольших количествах пептидазы, амилаза, нуклеаза, щелочная фосфатаза и другие ферменты. Образование и накопление их происходит в эпителиальных клетках, отторгающихся и подвергающихся распаду. Небольшого количества ферментов, выделяемых в толстые кишки, вполне достаточно для того, чтобы частично подвергнуть перевариванию пищевые вещества, поступившие из тонкой кишки. Е. С. Лондон указывал, что ферментовыдели-тельная способность толстого кишечника может в некоторой степени принимать участие в компенсации нарушенных функций и в вышележащих участках кишечника. В соке обычно содержатся в небольшом количестве фосфолипиды.
В толстых кишках совершаются движения трех типов: простые волны неодинаковой продолжительности в разных участках, причем и кишечное содержимое они не продвигают; более сильные и продолжительные, способны продвигать его; тонические волны, на которые накладываются фазные сокращения двух других типов. В связи с происходящим повышением гидростатического давления они могут стимулировать всасывание воды. Сокращения толстых кишок чередуются с периодами отсутствия моторики.
Рентгенологическими исследованиями выявлены: малые маятникообразные движения в проксимальных участках, способствующие в основном перемешиванию и разжижению содержимого; большие маятникообразные, участвующие в перемешивании содержимого и всасывании. Отдельные перистальтические сокращения мало эффективны в отношении продвижения содержимого вдоль кишок. Антиперистальтические движения способствуют перемешиванию и сгущению содержимого за счет всасывания преимущественно воды.
Продолжительность эвакуации, т.е. время, в течение которого происходит освобождение кишок от содержимого, у здорового человека достигает 24—36 ч. Согласно рентгенологическим данным, у человека контрастная масса через 3—3,5 ч после приема пищи начинает поступать в толстую кишку. Заполнение всей толстой кишки в среднем продолжается около 24 ч.
Толстый кишечник имеет как интрамуральную, так и экстрамуральную иннервацию. Рефлекторные влияния на моторику возможны с самых проксимальных участков пищеварительного тракта, она возбуждается при еде (Л. А. Семенюк) и осуществляется по принципу условного я безусловного рефлексов; рефлексы при прохождении пищи по пищеводу; с желудка на кишечник при раздражении и механо- и хеморецепторов. Рефлекторные влияния реализуются через блуждающие и чревные нервы с замыканием рефлекторной дуги в центральной нервной системе. Кроме того, доказана возможность распространения возбуждения с верхних отделов кишечника по стенкам кишечной трубки (П. Г. Богач). Раздражение стенки толстой кишки стимулирует ее моторику, при этом интенсивность моторики зависит от количества химуса, его консистенции, содержания в нем клетчатки, фекальных масс, газов, продуктов брожения и пр. Рефлекторное торможение моторики вышележащих отделов тонкого кишечника и толстого наблюдается с ампулы прямой кишки при наполнении ее каловыми массами (С. С. Полтырев).
В регулировании моторики толстой кишки участвуют и гуморальные факторы. В частности, серотонин тормозит моторику толстой кишки и стимулирует моторику тонкой кишки. Тормозящее действие оказывает адреналин, усиливающее — кортизон. Снижает двигательную активность глюкагон.
В толстом кишечнике содержится большое количество бактерий, вызывающих процессы брожения углеводов и гниения белков. Под влиянием бактерий расщепляется клетчатка. Поэтому роль бактерий особенно важна в пищеварении травоядных животных, длина толстого кишечника которых значительно больше, чем у плотоядных. У жвачных животных остатки корма в толстом кишечнике задерживаются в течение нескольких суток.
Под влиянием действия бактерий на нерасщепленные белки и продукты их распада в толстом кишечнике образуется ряд вредных для организма ядовитых веществ — индол, скатол, фенол, крезол и др., которые, всасываясь в кишечнике, могут вызвать интоксикацию. И. И.Мечников впервые обратил внимание на процессы гниения в толстых кишках как на источник интоксикации организма. Однако ядовитые вещества, образующиеся в толстом кишечнике в процессе гниения, всасываясь в кровь и по воротной вене поступая в печень, подвергаются в ней обезвреживанию. Следовательно, печень в этом отношении играет роль барьера, предохраняющего организм от действия токсических веществ.
Микрофлора кишечника изменчива в связи с характером питания, причем один вид микробов может вытесняться другим. Так, например, микробы молочнокислого брожения могут вытеснять гнилостные микробы (И. И. Мечников).
Кишечная микрофлора способна синтезировать ряд биологически активных соединений, как, например, витамин К и некоторые витамины группы В, с последующим всасыванием.
Вследствие всасывания воды содержимое толстого кишечника приобретает густую консистенцию. В формировании кала имеют значение плотные частицы кишечного сока, слизь, которые как бы склеивают непереваренные остатки пищи. Кал содержит остатки непереваренной пищи, растительную клетчатку, небольшое количество белков, жиров, углеводов, слизь, эпителий слизистой оболочки желудочно-кишечного канала, холестерин, продукты изменения желчных пигментов, придающие ему характерную желто-коричневую окраску, нерастворимые соли и различные бактерии. Последние составляют иногда 30—50% кала. Количество кала, выделяемого за сутки, зависит от количества поступившей в пищеварительный канал пищи, качества ее (растительная или животная), степени переваривания и усвоения пищевых веществ.
Удаление каловых масс осуществляется с помощью акта дефекации, представляющего процесс опорожнения нижних отделов толстого кишечника. Осуществляется он при помощи рефлекторного механизма. При наполнении ампулы прямой кишки калом происходит раздражение барорецепторов ее стенок. Возникающие при этом импульсы направляются в центр дефекации, расположенный в поясничной и крестцовой частях спинного мозга. Оттуда по центробежным нервам импульсы идут к гладкой мускулатуре толстого кишечника, мышцам внутреннего и наружного сфинктеров прямой кишки.
При этом происходит расслабление сфинктеров при одновременном резком сокращении мускулатуры толстого кишечника и прямой кишки. Акту дефекации способствуют сокращения мускулатуры брюшной стенки, диафрагмы и мышц, поднимающих анальный сфинктер.
Сфинктеры прямой кишки вне акта дефекации закрыты вследствие тонического сокращения их мускулатуры, причем сила сокращения мышц наружного сфинктера равна 1/2 или 1/3 общей силы сокращений мускулатуры сфинктеров.
При перерезке спинного мозга выше центра дефекации наблюдается непроизвольная дефекация. При разрушении центра дефекации или при перерезке спинного мозга ниже этого центра мускулатура анального сфинктера расслабляется и каловые массы выделяются наружу без задержки. Резкое и довольно длительное расслабление анального сфинктера происходит и при сильных эмоциях, например при страхе. В этих случаях наблюдается непроизвольная дефекация («медвежья болезнь»).
Спинномозговой центр дефекации подчинен в своей деятельности центру, расположенному в зрительных буграх промежуточного мозга, а этот в свою очередь подчинен коре больших полушарий.
Моторные нервы к сфинктерам кишки идут от передних крестцовых корешков спинного мозга в составе п. pelvicus, являющегося парасимпатическим нервом. Симпатические волокна подходят к сфинктерам через передние корешки поясничных нервов и нижний брыжеечный симпатический узел.
Глава 10
ПИЩЕВАРЕНИЕ ПРИ БЕРЕМЕННОСТИ И ЛАКТАЦИИ
На собаках с изолированным желудочком по Павлову и с гастроэзофаготомией было установлено, что во время беременности происходит резкое угнетение секреторной деятельности желудка, особенно в период слож-порефлекторной фазы.
Во время нормальной беременности у женщин наблюдаются значительные изменения в составе желудочного сока, главным образом за счет снижения кислотности и уменьшения переваривающей силы. Наиболее резкие изменения секреторного процесса отмечаются в первые месяцы беременности и за несколько недель до родов.
Объектом экспериментальных исследований связи лактации и пищеварения, как правило, являются домашние животные, в частности козы.
Опыты И. И. Грачева показали, что при механическом раздражении соска у коз происходит рефлекторное отрыгивание содержимого рубца и начинается вторичное пережевывание корма. Реакция обычно наступает быстро — через 3—15 с. Отмечено, что жвачный рефлекс, возникающий при механическом раздражении соска, очень стойкий: за 5—6 ч производилось до 75 раздражений с интервалами 3—5 мин, и при каждом из них наступала жвачка. Важно отметить, что на механическое раздражение соска жвачная реакция появлялась даже у голодных животных (после 5—6-суточного голодания), но при этом длительность реакции у голодных и накормленных животных неодинаковая, что указывает на важное значение исходного функционального состояния пищеварительного аппарата (С. С. Полтырев, И. П. Салмин).
Доказательством наличия воспринимающей части рефлекторной дуги жвачного рефлекса с рецепторов соска и поверхности молочной железы служат опыты с новокаинизацией тканей соска и нижней части вымени козы. После проведенной инфильтрационной анестезии жвачный рефлекс полностью исчезал и восстановление его наступало только через 45—60 мин. Следовательно, поверхность молочной железы обладает периферическими чувствительными нервными окончаниями, которые трансформируют механическое раздражение в нервные импульсы; последние по афферентным путям передаются в центральную нервную систему, а затем по эфферентным путям к пищеварительному аппарату
Наиболее достоверное подтверждение рефлекторного механизма связи между молочной железой и органами пищеварения дали опыты с денервацией вымени. После перерезки всех нервов, подходящих к молочной железе, в том числе и кожных, не удавалось вызывать жвачный рефлекс механическим раздражением денервированного соска. С другого интактного соска рефлекс продолжал воспроизводиться безотказно. У недоящихся овец механическое раздражение соска не вызывало жвачного рефлекса, так как такое воздействие было неадекватным. Сосание же ягненка сопровождалось появлением характерного жвачного рефлекса. Высказанное предположение, что при длительном доении овец механическое раздражение соска может быть стимулом для возникновения жвачного рефлекса, подтверждено экспериментом.
Все это не оставляет сомнения в том, что между молочной железой и пищеварительным аппаратом существует тесная рефлекторная связь и, кроме того, акт сосания — стимул для усиления процессов пищеварения. Эту связь можно рассматривать как наследственно закрепленный рефлекс, вызванный внешними воздействиями у животных при акте сосания и доения.
И. И. Грачеву удалось установить наличие натурального условного рефлекса у коз на тактильное радраже-ние рецепторов молочной железы. Следовательно, тактильное раздражение — условный сигнал безусловного механического раздражения. В дальнейшем было показано, что у животных можно образовывать условные рефлексы на индифферентные раздражители: звонок, свет электрической лампочки и др., а также на обстановку опыта.
В опытах на полифистульных козах и кроликах было отмечено, что при раздражении рецепторов молочной железы происходит усиление сократительной деятельности всех отделов многокамерного желудка коз и желудка кроликов, при этом происходит усиление сокращений. После суточного голодания коз раздражение рецепторов соска стимулировало сокращения всех отделов сложного желудка. Даже на фоне атропинизации, обычно тормозящей сократительную функцию пищеварительного тракта, механическое раздражение соска способствовало восстановлению движений и как бы устраняло торможение, вызванное атропином. Установлено, что на основе безусловнорефлекторного влияния на двигательную деятельность желудка козы со стороны соска могут быть выработаны условные рефлексы. Обнаружена рефлекторная стимуляция не только двигательной деятельности сложного желудка, но и желчного пузыря. Больше того, механическое раздражение соска может усиливать секрецию желчи, стимулировать сокращения слепой кишки. Е. М. Федий отметил, что при доении у коров увеличивается скорость прохождения корма по желудочно-кишечному тракту.
По данным П. В. Полетаева, раздражение рецепторов молочной железы у крольчих усиливает развитие пищеварительного аппарата. К. Т. Ташенов показал, что с наступлением лактации у коз возрастает секреция поджелудочного сока в 1,5 раза и удерживается на этом уровне в течение первой половины лактационного периода. В дальнейшем, по мере снижения секреции молока, уменьшается количество желчи и поджелудочного сока, а при прекращении лактации уровень секреции их снижается до уровня, характерного для нелактиру-ющих животных. Важно отметить, что наряду с количественными изменениями сокоотделения изменяется и содержание в соках плотных веществ, в том числе и электролитов. В поджелудочном соке повышается активность липазы.
Проводить аналогию между лактацией и пищеварением у животных и человека нельзя; в частности, нет оснований отрицать, что и у женщин существует функциональная связь между молочными железами и органами пищеварения, осуществляемая рефлекторным путем. Если учесть, что для образования грудного молока в количествах, необходимых для вскармливания ребенка, женщине требуется повышенное питание, то использование организмом потребляемых продуктов возможно лишь при усиленной деятельности пищеварительных органов и прежде всего главных пищеварительных желез.
Надо полагать, что еще до начала лактации между пищевым центром и центром лактации устанавливается определенная функциональная связь, от которой зависит в дальнейшем процесс молокообразования и повышенная способность пищеварительного тракта обеспечивать переваривание большего, чем обычно, количества потребляемой пищи лактирующей женщиной.
Глава 11
ПИЩЕВАРЕНИЕ ПРИ МЫШЕЧНОЙ ДЕЯТЕЛЬНОСТИ
Физическая работа, являясь для организма одним из необходимых компонентов жизнедеятельности, играет в филогенезе важную роль, а на определенных его этапах— решающую. Установлено, что двигательная активность активизирует не только деятельность опорно-двигательного аппарата, но и воздействует на различные функциональные системы организма и обмен веществ. Механизм воздействия на внутренние органы и обменные процессы во многом объясняет теория моторно-висце-ральных рефлексов, созданная М. Р. Могендовичем. Согласно ей ведущая роль в осуществлении влияний со стороны опорно-двигательного аппарата на различные функции принадлежит нервной системе. Однако в последние годы установлено участие в этих влияниях и ряда желез внутренней секреции, в особенности гипофиз-адреналовой системы, щитовидной железы.
Вопрос о влиянии мышечной работы на функции пищеварительного аппарата давно привлекал внимание физиологов и клиницистов.
До того как классический павловский фистульный метод нашел применение в физиологической науке, неоднократно делались попытки выяснить влияние мышечных напряжений на работу органов пищеварения. Так, в конце XVIII в. Матвей Пекен указывал, что движение подкрепляет желудок, помогает перевариванию пищи. В XIX в. появились работы посвященные изучению влияния мышечной работы на процессы пищеварения. Исследователи вели наблюдения за продолжительностью пребывания пищи или воды в желудке собак, находившихся либо в состоянии относительного покоя, либо совершавших бег. Однако из-за несовершенства методических приемов полученные результаты были противоречивыми. Одни утверждали, что под влиянием физической нагрузки деятельность желудка ухудшается, другие отмечали, что пищеварение не зависит от выполнения животным мышечной работы, третьи находили усиление сокоотделения.
С введением в практику экспериментальных исследований фистульного метода стало возможным получать достоверные данные о влияниях опорно-двигательного аппарата на различные функции пищеварительной системы и механизмах этих влияний. На фистульных собаках в условиях хронического эксперимента изучалось состояние большинства пищеварительных функций при двух основных видах мышечной деятельности: динамической (циклической) и статической. При этом учитывалось, что статические усилия по сравнению с динамическими более утомительны (А. Н. Крестовников).
Влияние мышечной деятельности на секреторную и экскреторную функции главных пищеварительных желез
Влияние мышечной деятельноти на работу слюнных желез у собак было изучено недостаточно. Только в условиях хронического эксперимента на собаках было проведено обстоятельное исследование слюноотделения при беге в третбане и установлено, что во время движения животного со скоростью 3 км/ч безусловная реакция слюнных желез угнетается. При движении со скоростью 5—6 км/ч было еще больше выражено снижение слюноотделения.
Немало работ экспериментального характера посвящено изучению работы желудочных желез при мышечной деятельности. Однако первой работой, выполненной в лаборатории И. П. Павлова в 1905 г. с целью изучения влияния мышечной работы на деятельность желудка, была работа И. С. Кадыгробова. В его исследованиях нагрузкой служила перевозка животным тележки с грузом около 5 и 10 кг. За шестичасовой опыт собака проходила с грузом расстояние в 7,5 км. Работа чаще выполнялась до кормления, а в некоторых опытах сразу после еды. Было установлено, что мышечная работа, выполнявшаяся животным до кормления, не оказывала заметного влияния на желудочную секрецию. Аналогичная ей физическая работа, выполненная непосредственно после кормления, тормозила сокоотделение в желудке в первые часы секреторного периода и повышала интенсивность секреции желудочного сока в последующие часы. Представляет интерес указание автора на то, что у животного по мере адаптации к физической работе изменения в деятельности желудка постепенно сглаживались, достигая затем нормы. У собаки, выполнявшей мышечную работу, эвакуация пищи из желудка в двенадцатиперстную кишку происходила ускоренно.
В дальнейшем многие исследователи наблюдали у собак торможение желудочной секреции при перевозке груза массой 3 кг.
Уменьшалось количество отделяемого на мясо и либиховский экстракт желудочного сока и снижалась его кислотность. При прогулке животного без груза в течение 2—6 ч также происходило снижение сокоотделения, но в меньшей мере. Было выяснено, что секреция желудочных желез у собак при выполнении легкой и тяжелой работы изменяется по-разному: легкая работа (перевозка груза массой 20 кг в течение 15 мин) вызывала усиление желудочной секреции и переваривающей способности желудочного сока на мясо и молоко, по при этом на молоко секреторный период не сокращался, а на мясо и хлеб уменьшался. Более тяжелая работа (перевозка груза массой 25 кг в течение 25—30 мин) вызывала снижение количества, кислотности и переваривающей силы желудочного сока на мясо и хлеб, на молоко же переваривающая сила была высокой.
В опытах, проведенных на трех фистульных собаках, которые получали мышечную нагрузку в виде бега в третбане в течение 30 минут (за этот период животное пробегало 6,4 км), было установлено, что мышечная работа, выполненная после кормления, полностью уничтожает рефлекторную фазу желудочной секреции и значительно тормозит химическую. Мышечная работа, производимая перед кормлением, заметного влияния на динамику желудочной секреции не оказывает.
Экспериментально установлено, что изменения секреторной функции желудка находятся в прямой зависимости от интенсивности мышечной работы и ее продолжительности (Т. И. Свистун). Так, движения собак в третбане со скоростью 3,0—3,5 км/ч вскоре после кормления вызывали угнетение желудочной секреции в первую фазу секреторного процесса, во вторую фазу она не изменялась. При скорости 5—8 км/ч угнетение желез желудка проявлялось в обе фазы. Мышечная нагрузка, даваемая за 1 ч до приема корма, не только не угнетала, но в ряде случаев даже повышала желудочную секрецию при указанных скоростях бега.
Исследованиями было показано, что под влиянием статических нагрузок, заключавшихся в удержании собакой на спине груза, равного половине массы тела животного (5 мин работы чередовались трехминутными периодами покоя), функция желудочных желез существенно изменялась: наблюдалось угнетение сокоотделения в случаях, когда нагрузка применялась сразу же после кормления, на фоне начавшейся желудочной секреции, если она предшествовала кормлению, и заканчивалась за 30 мин до него. Торможение проявлялось как в первую, так и во вторую фазу секреторного процесса. После снятия груза количество сока возрастало и повышалась его кислотность. Проведенное сопоставление влияния десятиминутной статической (0,5 кг на 1 кг массы тела животного) и динамической нагрузок (трехминутный бег в третбане со скоростью 7—15 км/ч) показало, что у животного наблюдалось торможение секреции желудочного сока.
Учитывая, что в практической деятельности человека могут иметь место перегрузки, и в том числе статические, необходимо было изучить в условиях хронического эксперимента влияние статических нагрузок значительной интенсивности и длительности на основные функции желудка и раскрыть механизм их. С этой целью Т. Н. Хрусталевой были проведены опыты на фистульных собаках.
При ежедневном применении статических нагрузок в течение 2 ч на протяжении от одного до четырех месяцев неизменно наблюдалось торможение секреции желудочного сока в обе фазы секреторного процесса, причем с увеличением нагрузки с 50 до 150% от массы тела животного степень выраженности угнетения возрастала. Одним из проявлений секреторных расстройств являлась также диссоциация между разными компонентами секреторного процесса (количество сока, его кислотность и переваривающая сила). Было установлено, что предшествующее двухнедельное применение статической нагрузки, равной 50% от массы тела, несколько ослабляло отрицательное влияние 100 и 150%-ной нагрузок, но полностью его не устраняло.
В результате длительного применения больших статических нагрузок, как показали гистологические исследования, в желудке развивались патоморфологичес-кие изменения дистрофического характера. Возможно, этим было обусловлено то, что измененная желудочная секреция стойко удерживалась и после прекращения опытов с нагрузками. В некоторых случаях желудочная секреция не достигала фоновых данных и спустя гри месяца после прекращения опытов.
Л. А. Шитовым было показано, что статические нагрузки (равные 100 и 150% от массы тела животного), использованные в опытах с изучением их влияния на работу желудочных желез, вызывали характерные признаки перенапряжения сердца. Следовательно, статическое перенапряжение обусловливает функционально-морфологические изменения не только в желудке, но и в сердце.
Функция поджелудочной железы исследовалась у собак при выполнении ими динамической работы. Было выяснено, что после тяжелой работы по перевозке груза в 20 кг в течение 1,5 ч у животного происходила задержка секреции поджелудочного сока. Это обнаруживалось в первые 30 мин после еды хлеба и мяса, а после еды масла уменьшалось общее количество сока.
С
одержание ферментов понижалось после еды хлеба и молока и не изменялось после мяса. Легкая работа (перевозка того же груза в течение 30 мин) не вызывала изменений сокоотделения при еде молока и мяса, при еде хлеба секреция повышалась в первый час, а при еде масла — за весь секреторный период.
Наблюдавшееся торможение могло быть обусловлено понижением сокоотделения в желудке и соответственно меньшим поступлением в кишечник кислого желудочного сока — возбудителя поджелудочной секреции. Надо отметить, что не исключалось и непосредственное тормозящее влияние мышечного напряжения на внешнесекре-торную функцию поджелудочной железы.
У собак с хронической фистулой протока поджелудочной железы наблюдалось при беге в третбане со скоростью 3—3,5 км/ч усиление поджелудочной секреции, несмотря на то, что при той же нагрузке желудочная секреция снижалась. Наряду с увеличением количества отделяемого сока повышались его щелочные резервы и липолитическая активность. Активность амилазы и трипсина не изменялась. Увеличение скорости до 5,5—6 км/ч приводило, наоборот, к угнетению секреции на пищевые вещества при одновременном снижении щелочных резервов в соке. Вместе с тем концентрация ферментов не изменялась.
Влияние мышечной деятельности на желчеотделение у собак изучено недостаточно. Выяснено, что мышечная работа, произведенная сразу после еды, вызывает у собак снижение желчеотделения, причем установлена зависимость степени выраженности торможения отделения желчи от скорости бега: чем больше скорость, тем сильнее выражено угнетение. Исследования показали, что в отделяемой желчи снижалось содержание кислот, пигментов и плотного остатка преимущественно за счет органических веществ. Выход желчи в двенадцатиперстную кишку на основные пищевые вещества во время движения собаки в третбане изменялся в зависимости от характера пищи.
Влияние мышечной деятельности па секреторную функцию кишечных желез также мало изучено. Известно лишь, что в отличие от желудочных желез при движении собак в третбане происходит усиление сокоотделения, вызываемого как механическим раздражением изолированной петли кишки, так и химическим. Усиление секреции отмечается у голодного (периодическая голодная секреция) животного и накормленного. При статических нагрузках наряду с усилением секреции происходит повышение ферментативной активности кишечного сока.
О вопросе экскреторной функции желудочных желез известно мало. При непродолжительном, но интенсивном беге собаки в третбане содержание в желудочном соке продуктов азотистого обмена (аммиака, мочевины) увеличивается. Усиление экскреции наблюдается и в случае применения динамических нагрузок через час и сразу после кормления. Повышенное содержание мочевины в желудочном соке обнаружено у собак и при статических напряжениях. Существует некоторая зависимость между изменениями секреторной и экскреторной деятельностью пищеварительных желез при выполнении мышечных нагрузок динамического характера.
Следовательно, железы желудка (в условиях мышечной работы) выделяют продукты азотистого обмена более интенсивно, чем в состоянии покоя. Этим самым они до известной степени облегчают экскреторную функцию почек.
Влияние мышечной деятельности на двигательную функцию желудочно-кишечного тракта
Динамическая работа оказывает существенное влияние на двигательную функцию желудка. Известно, что ограничение двигательной активности ослабляет секреторную и двигательную функции пищеварительного аппарата. Так, М. Р. Могендовичем с сотрудниками изучалась периодическая моторика желудка в условиях иммобилизации конечности у собаки и было установлено, что наложенная на 6—13 дней гипсовая повязка с первого же дня вызвала изменения в периодической деятельности желудка. После снятия повязки голодная периодическая деятельность желудка постепенно возвращалась к исходному состоянию. Исследования показали, что чем длительнее продолжалась иммобилизация конечности, тем меньшими функциональными сдвигами она сопровождалась. Это указывает на происходившую адаптацию к длительному воздействию иммобилизации.
В опытах на собаках было показано, что бег животного в топчаке сразу после введения бульона угнетал моторику желудка. Если же бег происходил через 5— 8 мин после введения пищи, то тормозящее действие заметно ослаблялось.
Статическое напряжение, вызываемое грузом 4— 12 кг, наложенным на спину на 10—20 мин, вызывало у собак угнетение периодики желудка. Однако иногда происходило усиление моторики. У собак при беге в третбане со скоростью 3,5—5,5 км/ч период работы укорачивался за счет уменьшения количества отдельных сокращений; при быстром передвижении животного (до 8 км/ч) наступали более резкие изменения периодики. Тонус желудка при обеих скоростях бега повышался.
При применении статической нагрузки (равной 50% от массы тела) в течение 1 ч с перерывами, приуроченной к началу периода голодных сокращений, происходило угнетение их, которое сохранялось и во втором периоде сокращений, когда груз был снят. Лишь в отдельных опытах первоначальное торможение сменялось усилением сокращений. Десятиминутное удержание груза (равного массе тела животного), примененное в начале периода голодных сокращений, угнетало их, но после прекращения действия статической нагрузки они появлялись вновь. Менее выраженное угнетение вызывало часовое применение статической нагрузки. В опытах с динамической нагрузкой (бег в третбане со скоростью 3—3,5 и 5,5—7,5 км/ч) также наблюдалось торможение периодической деятельности желудка.
При статических нагрузках большей продолжительности (в течение 2 ч) и большей интенсивности (100 и 150% от массы тела) у собак наблюдается извращение кривых голодных сокращений желудка, изменения продолжительности периодов работы и относительного покоя. Изменения тонуса стенки желудка под влиянием этих нагрузок отражается и на сроках опорожнения желудка. Показано, что бег в третбане тормозит эвакуацию содержимого из желудка в двенадцатиперстную кишку. Вместе с тем эффект зависит не только от величины нагрузки и скармливаемой пищи, но и от времени начала бега. Если он предшествовал кормлению, то задержка эвакуации была менее длительная, чем после кормления. Изменения эвакуации вызывали и статические нагрузки. При нагрузке в 2—4 кг опорожнение желудка происходило ускоренно, а при 10 кг в начале эвакуация задерживалась, а затем ускорялась. Следует отметить, что между увеличением нагрузки и усилением эвакуации пропорциональная зависимость отсутствует.
Статические нагрузки, равные 100 и 150% от массы тела, замедляют эвакуацию из желудка 2%-ного раствора крахмала и 0,25%-ного раствора соляной кислоты, однако в тех случаях, когда применению этих нагрузок предшествовала тренировка нагрузками, равными 50% от массы тела, задержка эвакуации была менее выражена. Этот факт указывает на возможность проявления адаптации к большим нагрузкам.
В 70-х годах XX в. было показано, что под влиянием больших статических нагрузок (100 и 150% от массы тела) изменяется двигательная деятельность не только желудка, но и тонкого кишечника. Как правило, у собак отмечается торможение моторики тонкой кишки. Было высказано предположение, что в условиях усиленного отделения кишечного сока, вызываемого действием больших нагрузок, и повышенного содержания в нем кишечных ферментов замедление моторики можно рассматривать как приспособительную реакцию, способствующую более полному всасыванию в тонкой кишке продуктов переваривания пищи.
Таким образом, мышечное напряжение статического и динамического характера оказывает существенное влияние на двигательную деятельность желудка и тонкой кишки как у голодного животного, так и накормленного. Под влиянием мышечных нагрузок изменяется не только голодная периодика, эвакуаторная деятельность желудка, но и процесс всасывания в кишечнике.
Е. С. Лондон и Н. Кочнева наблюдали снижение всасывания в кишечнике аминокислот и полипептидов у ангиостомированных собак, перетаскивающих тележку в течение 30 мин. Изменения скорости всасывания в желудке и кишечнике у собак отмечали многие исследователя. При беге в третбане со скоростью 5,5 км/ч в течение 30 мин у собак усиливалось всасывание в тонкой кишке глюкозы; при беге с той же скоростью в течение 2 ч всасывание угнеталось. Следовательно, характер изменения интенсивности всасывания зависит от величины мышечного напряжения.
Усиление всасывания жира (0,5 мл меченого 131I триолеина в оливковом масле) было установлено у крыс при плавании в течение 40—180 мин.
Под влиянием статической нагрузки, равной 0,5 кг на 1 кг массы тела собак и продолжительности ее действия в течение 1 ч с интервалами (10 мин нагрузка и 5 мин отдых), было установлено усиление всасывания глицина и снижение всасывания глюкозы в желудке. На всасывание воды данная нагрузка влияния не оказывала. С увеличением ее (0,75—1,5 кг/кг) происходило угнетение всасывания глицина и глюкозы, но при этом всасывание воды по-прежнему не изменялось. Более резко был выражен эффект при беге собак со скоростью 13,5 км/ч. Однако со снижением скорости до 7,5 км/ч стимулирующий эффект снижался. Отмечено, что повышение всасывания имело место только во время бега, а спустя 1 ч после прекращения его интенсивность возвращалась к исходному уровню.
Статические нагрузки, равные 50—75% от массы тела животного, не оказывали влияния на скорость всасывания в тонкой кишке глицина и 1%-ного раствора поваренной соли; при большей нагрузке (100—150% ОТ массы тела) уровень резорбции резко снижался. При статических нагрузках, равных 50 и 75% от массы тела, несколько усиливалась резорбция в тонкой кишке глюкозы и воды, а при большой нагрузке (100 и 150%) происходило выраженное угнетение всасывания этих веществ. Эффект удерживался некоторое время и после снятия груза.
Подтверждением того, что реакция со стороны всасывательного аппарата неодинакова в зависимости от применяемых веществ, было следующее наблюдение. Во время движения животного в третбане со скоростью 3 км/ч увеличивалось всасывание в тонкой кишке собак воды из растворов глюкозы и аминокислот, возрастало всасывание и самой глюкозы. Что же касалось всасывания самих аминокислот, то оно или не изменялось, или даже угнеталось.
Механизмы влияния мышечных нагрузок на функции желудочно-кишечного тракта. Предположения относительно механизма влияний физических напряжений высказывались давно, но далеко не все получили в дальнейшем подтверждение. Так, И. С. Кадыгробов связывал увеличение секреции в 4—5-е часы опыта с нагрузкой, с повышением у животного аппетита, а уменьшение секреции в желудке он объяснял обеднением организма водой в связи с усиленным потреблением ее работающими мышцами. Изменения желудочной секреции он пытался объяснить сдвигами в эвакуации содержимого из желудка в двенадцатиперстную кишку. Видя несостоятельность подобных предположений, И. П. Павлов в 1905 г. указал па то, что вопрос о причине извращения отделения желудочного сока при мускульной работе остается невыясненным.
Высказывалось предположение, что изменения секреторной деятельности желудка зависят oт накопления и организме при мышечной работе молочной и угольной кислот, изменениями температуры тела и пр. Некоторые придавали большое значение происходящей анемизации внутренних органов вследствие усиленного кровоснабжения работающих мышц. Конечно, нельзя отрицать влияния указанных факторов па работу желудочных желез при мышечной деятельности, но нельзя также их переоценивать. Как показали исследования последних лет, главная роль в осуществлении влияний мышечных напряжений на работу органов пищеварения принадлежит нервной системе и железам внутренней секреции.
С тех пор, когда было установлено наличие в скелетных мышцах, сухожилиях, связках, суставах проприоре-цепторов, возбуждаемых во время двигательных актов, стал понятен рефлекторный механизм влияний мышечной деятельности на различные стороны жизнедеятельности организма. Как выяснилось, возникающие в проприоре-цепторах импульсы при осуществлении двигательных актов направляются затем по афферентным путям в центральную нервную систему, в том числе и в кору головного мозга, а оттуда после соответствующего анализа они посылаются в различные органы и системы организма. Эти рефлексы были названы М. Р. Могендовичем моторно-висцеральными.
Физическая работа оказывает влияние на железы желудка через вагус и симпатикус. При мышечных нагрузках тормозные влияния передаются в основном по вагу-су, а стимулирующие — по симпатикусу (Т. И. Свистун). Это, в частности, обусловливает угнетение первой и усиление второй фазы секреции желудочного сока при беге животного в третбане с небольшой скоростью (3—3,5 км/ч). Однако в механизме влияния мышечной работы ведущая роль принадлежит коре головного мозга. Установлено, что физическая работа изменяет функциональное состояние мозговой коры.
Так, при беге в третбане с указанной выше скоростью у собак происходит усиление раздражительного процесса и ослабление тормозного. Через 4—6 месяцев тормозной процесс восстанавливается, но при этом возбудимость корковых клеток во время бега остается в течение продолжительного времени повышенной. При большей скорости бега (5—6 км/ч) возбудимость коры головного мозга понижается.
Произведенный Т. Н. Хрусталевой анализ механизма влияний больших статических нагрузок на работу желудочных желез показал, что выключение парасимпатической иннервации желудка (собаки с гей-денгайновским желудочком) не устраняет возможности проявления тормозного влияния статических нагрузок, равных 100 и 150% от массы тела животного. Вместе с тем тормозное влияние менее выражено, чем у собак с павловским желудочком при тех же нагрузках. Следовательно, тормозные влияния осуществляются на железы желудка в основном через вагус. Интересные данные получены в опытах на собаках . с полностью де-нервированным желудочком по Грегори. Опыты показали, что и после полной денервации изолированного желудочка большие статические нагрузки продолжают оказывать влияние на работу желудочных желез, однако тормозной эффект выражен слабее, а при нагрузке 150% от массы тела он не только отсутствовал, но наблюдалось даже его стимулирующее влияние. Все эти данные позволили предположить, что в денервированнои желудочке реакция желудочных желез осуществляется не рефлекторным путем, а гуморальным. Это предположение получило подтверждение в опытах на собаках с удаленными щитовидной железой и одним надпочечником и денерви-рованным другим надпочечником. Так, у собак с интакт-ной щитовидной железой статические нагрузки, равные 100 и 150% от массы тела, резко снижали сокоотделение в желудке в 1—4-е часы секреторного периода, у собак с удаленной щитовидной железой во время 1-го часа стояния с грузом торможение почти отсутствовало, а во время 2-го часа и особенно 3—4-го часов секреция значительно возрастала. Следовательно, после выключения щитовидной железы в основном устранялось тормозное влияние статических нагрузок на желудочную секрецию в первую фазу секреторного процесса. По-видимому, в развитии торможения желудочной секреции при больших статических нагрузках принимает участие щитовидная железа.
На собаках с одним денервированным и другим удаленным надпочечником большие нагрузки не только не проявляли обычного тормозящего влияния на железы желудка, но, наоборот, оказывали стимулирующее действие. Следовательно, в развитии торможения участвовала не только щитовидная железа, но и надпочечники.
Произведенный фармакологический анализ с использованием дигидроэрготоксина, карбахолина, бензогексония, аминазина, фенамина показал, что характер реакции желудочных желез на фоне измененного функционального состояния разных отделов центральной нервной системы был иным, чем у собак с ненарушенным функциональным состоянием нервной системы. Этот результат подчеркивает важное значение исходного состояния нервной системы в формировании реакции на действие больших статических нагрузок.
Большие статические нагрузки вызывали изменения кишечной секреции, причем не только в интактной изолированной петле тощей кишки, но и в денервированнои. В опытах на животных с двумя изолированными петлями кишок, с одной интактной и другой денервированной, было установлено однонаправленное изменение секреции— усиление сокоотделения. Отличие состояло лишь в том, что из денервированной петли увеличение секреции было менее выраженным, чем из интактной. Такое различие в реакции можно объяснить тем, что на денервиро-ванную петлю кишки статические нагрузки действуют гуморальным путем, а, как известно, денервированные ткани обладают повышенной чувствительностью к химическим агентам, в том числе и гормонам.
Интересные данные получены в опытах на собаках с применением индерала — блокатора α-адренорецепторов. Индерал вызывал торможение секреторной функции желудка на пищевые вещества при нормальном состоянии желудочной секреции. На фоне же низких исходных показателей желудочной секреции и повышенной пищевой возбудимости эффект был противоположным. Этот факт еще раз подчеркивает значение исходного состояния в реакции на то или иное воздействие.
Участие нервной системы в осуществлении влияния мышечных нагрузок на всасывательную деятельность тонкой кишки было выявлено в специально проведенных опытах. При статической нагрузке, равной 100% от массы тела, у собак наблюдалось угнетение всасывания глюкозы в тонкой кишке в течение 34 дней после перерезки больших чревных нервов. Этот факт указывает на то, что тормозное влияние на кишечные железы осуществляется через блуждающие нервы. Двусторонняя перерезка чревных нервов устраняла стимулирующее влияние бега в третбане на процесс всасывания в желудке. Такой же эффект наблюдался и при блокаде вегетативных ганглиев тетамоном U. Поддиафрагмальная перерезка вагусов или перерезка серозно-мышечного мостика между большим и маленьким желудочками не устраняли стимулирующего влияния бега на всасывание в изолированном желудочке. Неодинаковое влияние бег оказывал на всасывание в желудке у собак с измененнным бромом и кофеином функциональным состоянием: при бромировании всасывание в покое снижалось, а во время бега — нормализовалось; на фоне применения кофеи-па бег угнетал всасывание в желудке глицина и глюкозы. Выяснено, что задержка эвакуации из желудка в двенадцатиперстную кишку при движении животного обусловлена усилением кислотного рефлекса. Однако оно может быть обусловлено и рефлекторным повышением тонуса гладких мышц желудка и двенадцатиперстной кишки.
Об участии коры головного мозга в осуществлении влияний мышечных нагрузок на органы пищеварения свидетельствуют следующие данные: в процессе многократного проведения опытов со статическими нагрузками у собак вырабатывался условный рефлекс на обстановку опыта. При повторных применениях статических нагрузок возникало условнорефлекторное торможение голодных сокращений.
Вопрос о влиянии мышечной работы на функции органов пищеварения у человека изучен недостаточно.
В опытах, проведенных па человеке после длительной ходьбы или упражнений со штангой, было выявлено угнетение слюноотделения на пищевые раздражители. После работы снижался рН слюны.
Исследования, проведенные па 18 здоровых людях с целью выяснения влияния статической и динамической работы на безусловнорефлекторное слюноотделение, показали, что под влиянием работы руки тормозится слюноотделение в среднем на 18—22%, в то время как общая статическая нагрузка (удержание груза массой 8 кг за плечами) стимулировала отделение слюны, причем в среднем па 22%. По мнению исследователей, противоположный эффект зависел от того, что напряженно работающая группа мышц руки индуцирует торможение на центр слюноотделения, тогда как статическое напряжение большого количества мышц, поддерживающих акт стояния с небольшим грузом, вызывает иррадиацию возбуждения, распространяющегося и на центр слюноотделения (М. Р. Могендович и др.).
Изучение функций желудка у человека при выполнении мышечной деятельности привлекло внимание многих исследователей. Во всех работах отмечается, что мышечная деятельность изменяет функциональное состояние желудочных желез, но при этом одни исследователи указывают на ее угнетающее влияние на секрецию желудочного сока, другие — на стимулирующее, а третьи — угнетающее и стимулирующее влияния. Различный эффект, который наблюдали исследователи даже при действии одного и того же мышечного напряжения, мог зависеть от ряда причин и, в частности, от неоднотиппости методик исследования, небольшого числа испытуемых и определения чаще всего в составе желудочного сока только соляной кислоты.
Особый интерес представляет вопрос об особенностях функций желудка у людей, систематически занимающихся спортом. Уманский исследовал у 159 спортсменов кислотность и переваривающую способность желудочного сока и при этом установил, что средние показатели кислотообразующей функции желудка у спортсменов разных специальностей были в пределах нормы. Различий, связанных со спортивной специализацией, в экскреции пепсиногена с мочой не было обнаружено. Эти выводы носят сугубо ориентировочный характер, так как были использованы косвенные методы определения кислотообразующей и ферментовыделительной функций желудка.
Фракционным зондированием секреторную деятельность желудка у людей изучали многие исследователи. Так, Г. Н. Пропастин у большинства из 26 спортсменов выявил более высокие показатели кислотообразующей и ферментовыделительной функций желудка, чем у здоровых людей, но не занимающихся спортом. Однако полученные цифры не выходят за пределы нормы. Повышенные показатели, по мнению автора, — следствие систематического применения физических нагрузок. Обнаружены существенные различия в реакции секреторного аппарата желудка на нагрузку у спортсменов и лиц, не занимающихся спортом. После 20—25-минутной гигиенической гимнастики у спортсменов в обе фазы секреторного процесса изменения в кислотообразующей и ферментовыделительной функциях отсутствовали, в то время как у лиц, не занимающихся спортом, было отмечено угнетение, причем преимущественно в первую фазу секреторного процесса. Бег в умеренном темпе в течение 40—60 мин у незанимавшихся спортом снижал в обе фазы объем выделяемого сока, а также качественные его показатели: у спортсменов, несмотря на то, что за этот период времени они пробегали значительно большую дистанцию (10—15 км вместо 8 км), угнетение имело место только в первую фазу. Г. Н. Пропастин вполне обоснованно объясняет различие в реакции желудочных желез на нагрузку разным уровнем адаптации испытуемых к мышечному напряжению.
А. А. Плешаковым исследовались особенности желудочной секреции у 423 человек, из них 390 были спортсменами в возрасте 18—22 лет.
В результате проведенных исследований им установлено, что занятия спортом изменяют секреторную деятельность желудочных желез, направленность и уровень сдвигов зависят от специализации, квалификации, стажа и возраста спортсменов. Различия в объеме секрета, содержании в нем соляной кислоты, хлоридов, пепсиногена, натрия и калия были обнаружены как при исследовании в покое, так и при мышечной деятельности (дозированная нагрузка) и выявлены в базальной и в меньшей мере в стимулированной секреции. Наиболее выраженная гиперфункция наблюдалась у велосипедистов, лыжников, конькобежцев.
Одним из проявлений адаптации желудочных желез к мышечным нагрузкам является увеличение их устойчивости к ней, причем уровень ее растет вместе с ростом тренированности спортсменов. Установлено, что менее частым вариантом адаптации является гипофункция желудочных желез, при этом она характерна для спортсменов, организм которых часто подвергается обезвоживанию в результате потери массы (например, у борцов, боксеров, штангистов).
В механизме регуляции секреторной деятельности желез желудка в условиях мышечной деятельности участвуют нервные и гуморальные факторы. Фармакологическое выключение парасимпатических и симпатических влияний, усиление симпатических влияний в условиях дозированной нагрузки показали, что устойчивость секреторной функции желудка к мышечным напряжениям, ее стимуляция обеспечиваются комплексом парасимпатических и симпатических влияний. Парасимпатикус обеспечивает регуляцию базальной и стимулированной секреции, а симпатикус оказывает влияние преимущественно па стимулированную секрецию. Среди эндокринных желез важное значение имеет кора надпочечников.
Глава 12
ПИЩЕВАРЕНИЕ ПРИ ЭКСТРЕМАЛЬНЫХ ВОЗДЕЙСТВИЯХ
И ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ
РАССТРОЙСТВ СЕКРЕТОРНО-МОТОРНОЙ ФУНКЦИИ
ПИЩЕВАРИТЕЛЬНОЙ СИСТЕМЫ
При действии на организм различных экстремальных факторов нормальная деятельность пищеварительных желез, желудочно-кишечного тракта, механизмов управления и регуляции может надолго расстроиться. К таким факторам, действие которых на пищеварение подверглось лабораторному и отчасти клиническому изучению, относятся: высокая температура внешней среды, пониженное атмосферное давление (гипоксемия), ускорения, вибрация и невесомость, различного вида излучения (УВЧ, ультрафиолетовая радиация, ионизирующая радиация и сверхсильные звуковые раздражения).
Пребывание человека или животного в условиях высокой внешней температуры оказывает на пищеварение существенное влияние. В основном оно сводится к угнетению секреторной активности пищеварительных желез и снижению моторной деятельности желудочно-кишечного тракта. Это хорошо выявлено на примере желудочных желез (И. II. Разенков, Н. В. Данилов, А. С. Садыков). Если собаки находятся в течение 45 мин в тепловой камере при температуре 50° С, то после этого у них резко снижается количество желудочного сока на прием пищи, особенно в период сложнорефлекторной фазы секреции. Латентный период сокоотделения увеличивается, продолжительность секреции сокращается. Торможение секреторного процесса происходит не только в день пребывания животного в условиях высокой внешней температуры, но и в течение 10—14 дней. Пребывание животного в тепловой камере при температуре 40е С или совсем не оказывает влияния на ход секреторного процесса, или вызывает небольшие сдвиги в сторону увеличения или снижения секреции только лишь в день опыта; при температуре 30° С стимулирует работу желудочных желез.
Наблюдения показали, что пребывание человека в течение 30 мин при температуре 60—65° С и высокой относительной в'лажности (70—80%) резко тормозит секреторную деятельность желудочных желез. Таким образом, работа желудочных желез после 30—45-минутного пребывания организма в атмосфере при температуре 30° С стимулируется, а при температуре 50° С и выше резко угнетается.
Подобные изменения наблюдаются со стороны других пищеварительных желез. Механизм их возникновения обусловлен, как показали специальные опыты, не только обезвоживанием организма в связи с потерей влаги, но главным образом тормозящим влиянием на секреторные клетки пищеварительных желез со стороны высших мозговых центров. В механизме угнетения принимают участие и гормональные вещества. Зеленый чай значительно снижает эффект теплового угнетения пищеварительных желез.
Существенное влияние на пищеварение оказывает в гипоксемический фактор, действие которого развивается при понижении барометрического давления и соответственно уменьшении парциального давления кислорода воздуха, что бывает, когда человек высоко поднимается в горы, летит самолетом на высоте более 3000 м или совершает подъем на различные высоты без кислородной маски в специальной барокамере пониженного барометрического давления. Изучение этого вопроса проводилось главным образом на собаках с фистулами протоков пищеварительных желез, а также на людях с помощью метода зондирования желудка. Секреция слюны изучалась с помощью капсулы Лешли— Красногорского.
В условиях пониженного барометрического давления животный организм испытывает кислородное голодание (недостаток кислорода в крови). Развивающаяся гипок-семия и как следствие этого гипоксия (недостаток кислорода в клетках и тканях) —основная причина возникающих в организме расстройств и нарушений функций. Они касаются окислительных процессов, всех видов обмена веществ, особенно углеводного, жирового и белкового, значительные изменения возникают в мозговых центрах и эндокринных железах; глубокие сдвиги наблюдаются в секреции пищеварительных соков. Весь этот комплекс изменений, сопровождающихся у человека сильной головной болью, головокружением, резким по-бледнением кожи лица и тела, тошнотой и рвотой, адинамией, одышкой, частым сердцебиением и обморочным состоянием, называется горной или высотной болезнью и проходит, как только человек вернется на «землю» или подышит чистым кислородом.
В исследованиях исходили из того, что высотная болезнь Связана с падением парциального давления кислорода в альвеолярном воздухе, и высота 4—5 км считалась границей бескислородного полета, высота 6 км — «критический порог», за которым до 8 км находится «критическая зона», где симптомы высотной болезни выражены резко. От 6 до 8 км парциальное давление кислорода в альвеолах падает до 32,5% и насыщение артериальной крови кислородом с 96%, как в норме, до 64% и более. Зона выше 8 км называется «порогом смерти».
При подъеме в барокамере на высоту 4—8 км (экспозиция 5—6 ч) у собак резко снижается слюнная секреция, особенно на отвергаемые вещества (0,25%-нын раствор соляной кислоты); у людей при таких условиях (высота 4 км) также возникает угнетение слюноотделительной функции желез. После 9—10 подъемов частотой 2—3 раза в неделю и пребыванием на высоте 5—6 ч отмеченные изменения слюноотделения становятся слабо выраженными или совсем исчезают. Такая адаптация дает положительный эффект у некоторых собак уже после пяти «высотных» подъемов. Но встречаются собаки, адаптация у которых не наступает даже после 1,5-месячного ежедневного подъема. Такие же изменения высотных расстройств секреции слюны и колебания эффекта адаптации при частых подъемах отмечались и у человека. Возможно, это связано с типологическими особенностями нервной системы животных и человека. Установлено также участие симпатической иннервации.
Анализ полученных экспериментальных данных о влиянии пониженного барометрического давления на секреторную деятельность желудка у людей и собак позволил сделать заключение о том, что при действии этого фактора возникает резкое торможение желудочной секреции, и воздействия осуществляются главным образом через рефлекторные механизмы, гуморальные механизмы принимают меньшее участие. При этом было установлено, что при еде хлеба желудочная секреция бывает резко сниженной, а при еде мяса, как и на прием гематогена и алкоголя, секреция может быть нормальной. Желудочные железы, как и слюнные, довольно хорошо адаптируются к действию гипоксемического фактора.
Степень гипоксемии имеет значение для быстроты эвакуации пищи из желудка в кишечник в условиях пребывания в барокамере на «высоте». Торможение желудочной секреции, возникающее у человека и животных при подъеме в барокамере на высоту 4—5 км, снижается или значительно ослабляется, если проводить соответствующую тренировку.
При полетах на самолетах секреция желудочного сока у человека на высоте около 5 км снижается на 40—50% по сравнению с контрольными («наземными») опытами. В 2,5 раза по сравнению с нормой замедляется эвакуаторная функция желудка (Ю. Н. Успенский). Тормозятся и функции кишечных желез, если собак поднять в барокамере на «высоту» 6—8 км без кислорода. Последействие бывает в этих случаях продолжительным, до двух месяцев. После подъема на высоту до 4 км у собак в первые часы наземных опытов секреция кишечных желез бывает повышенной в 1,5 раза. Период последействия длится около 12 дней. Изменения качественного состава сока менее выражены и быстро выравниваются. Кратковременные полеты на самолете (10—30 мин) на высоте до 4 км не оказывают влияния на деятельность кишечных желез. Но если они проходят с «болтанкой», то возникает резкое торможение кишечной секреции.
Наблюдения, проведенные Г. К- Шлыгиным во время Эльбрусской экспедиции на собаке с хронической фистулой протока поджелудочной железы, показали, что длительное пребывание животного на высоте 2 200 м (30 дней) вызывает повышение секреторной активности желез и увеличение щелочности поджелудочного сока; при подъеме на 4 250 м секреция в первые дни тормозится, а затем снова возрастает; при этом изменяется тип кривой секреции.
В результате опытов, проведенных И. Т. Курциным на собаках с павловской фистулой протока поджелудочной железы в камере пониженного барометрического давления, было установлено, что пятичасовое пребывание на высоте до 3 км стимулирует, а выше — угнетает пищевую секрецию поджелудочной железы. Значительное подавление активности секреторных клеток возникает на высоте 5—6 км и резкое угнетение ее наступает на высоте 9 км.
Существенные изменения, как показали исследования И. П. Разенкова и его сотрудников, наступают на высоте свыше 3 км и в секреции желчи. Хотя пребывание на этих высотах было кратковременным (около 15 мин), тем не менее изменения секреторной реакции па пищевой раздражитель (мясо, хлеб, молоко) были не только в день опыта в барокамере, но сохранялись затем в течение двух-трех недель после подъема на средние и большие высоты. Во всех опытах изменения касались не только количественной, по и качественной стороны секреции, например амилолитических, иротео-литических и липолитических свойств поджелудочного сока. Подавление активности то касалось всех ферментов, то только отдельных, например снижение активности амилазы и трипсина и увеличение активности липазы.
При первом подъеме на 4,5 км пищевая секреция увеличивалась, на повторном подъеме четко выявилось торможение секреции, которое наблюдалось затем в течение двух недель в опытах «на земле». В дальнейшем явление торможения исчезло и наступила резко выраженная гиперсекреция. На таком фоне при подъеме на ту же высоту опять возникло торможение пищевой секреции, которое сохранилось и в последующих опытах, проводимых вне камеры. Тормозной эффект был выражен главным образом в сложнорефлекторной фазе, а гиперсекреторный — в нейрохимической.
Показано, что шестичасовое пребывание в барокамере при разрежении, соответствующем разрежению на высоте в 6—8 км, вызывает у собак значительное снижение спонтанной секреции желчи и изменение качественного ее состава (плотный остаток, желчные кислоты, билирубин). Эти тормозные явления, подобные изменениям деятельности слюнных, желудочных и кишечных желез, характеризуются довольно большой стойкостью и при повторных подъемах иногда проявляются в более сильном виде, чем в первые подъемы. У некоторых собак адаптация к высотным факторам доходит до нескольких недель. Опыты, проведенные па высотах 7 и 9 км, показали наличие резкого угнетения секреторном функции поджелудочной железы в обе фазы секреции п сильное подавление ферментообразования. Тормозной эффект сохранялся и в опытах, проводившихся «на земле» в последующие дни после подъема на эти высоты. Последействие в этой серии опытов отмечалось около трех недель, после чего прием пищи вызывал нормальную реакцию железы.
Таким образом при действии на организм высотного фактора было выяснено, что подъем на высоту до 2— 3 км не оказывает угнетающего влияния на пищеварение.
При комплексных воздействиях на организм трудно ожидать однонаправленного эффекта даже в том случае, если хорошо изучено действие каждого фактора в отдельности. Взять, к примеру, ультрафиолетовую радиацию, действие которой на организм в естественных условиях во время подъема в гору происходит вместе с действием высотного фактора. При изолированном действии ультрафиолетовая радиация вызывает увеличение секреции желчи. Гипоксемический фактор (подъ-ем в барокамере) при таких же условиях вызывает торможение желчеотделения. При сочетании действия этих двух факторов (подъем в гору) секреция желчи угнетается. Но если в условиях барокамеры животное перед подъемом на высоту подвергнуть ультрафиолетовому облучению, то гипоксемический эффект угнетения секреции желчи снимается. Следовательно, ультрафио: летовая радиация как бы повышает резистентность клеток и тканей организма к кислородному голоданию. Подобное влияние оказывает электрическое поле УВЧ на секреторную и экскреторную функции тонкого кишечника собак, подвергнутых действию УВЧ, а затем гипо-ксемического фактора.
Основные закономерности изменений деятельности пищеварительных желез, открытые и изученные в условиях барокамеры пониженного барометрического давления и полета на самолете, подтвердились и в высокогорных условиях во время специальных экспедиций на Эльбрус на высоту 2 км 200 м, 4 км 250 м, 4 км 800 м.
Полученные данные как в эксперименте на собаках, так и в наблюдениях на людях позволили заключить, что основным фактором, действующим на пищеварительные железы на горных высотах, является недостаток кислорода. В результате приспособляемости, приобретенной за два месяца пребывания на высотах Эльбруса, деятельность желудочных желез и желчеотделение не подвергаются существенным изменениям при последующих (через три месяца) испытаниях в барокамере и в опытах на самолете. Однако при многократных подъемах на большие высоты (8 км) и длительное пребывание там (5 ч) приспособляемость может быть нарушена. Результаты опытов позволили сделать важный вывод: пониженное барометрическое давление, соответствующее высоте 8 км (три дня по 5,5 ч), практически не влияет на усвояемость пищевых веществ при смешанном пищевом режиме.
Проблема радиационных поражений органов пищеварения, возникшая перед учеными в связи с широким использованием атомной энергии в различных отраслях народного хозяйства, в настоящее время стала одной из актуальных проблем современной физиологии и клиники. Лучевые поражения организма и связанные с ними нарушения пищеварения могут происходить при действии различных видов ионизирующих излучений, т. е. таких лучей, которые имеют меньшую длину волн, чем инфракрасные, видимые и ультрафиолетовые, и при прохождении через какую-либо поглощающую среду вызывают процесс ионизации, т. е. образование ионов из нейтральных атомов и молекул. В состав радиоактивных излучений входят α- и β-частицы, рентгеновы и γ-лучи, нейтроны (медленные и быстрые) и протоны. Внешние источники этих излучений: рентгеновская аппаратура, урановые рудники, обогатительные фабрики, ядерный реактор, «горячие» лаборатории для обработки ядерного горючего и получения изотопов и места применения их в научных исследованиях, сельском хозяйстве, промышленности, медицине, ветеринарии, взрыв атомной или водородной бомбы, а также общеприродный фон: воздух, почва, растения, вода и пища. Внутреннее облучение организма может происходить от α-, β- и γ-лучей, источниками которых служат радиоактивные вещества, поступившие в тело.
Все излучения можно разделить на три группы: 1) заряженные частицы, 2) нейтроны, 3) электромагнитные излучения. Биологическое действие их в принципе одинаково, различна лишь сила действия.
В рентгенологии и радиологии за единицу дозы рентгеновых и у-лучей принят рентген. Для оценки других видов радиоактивных излучений предложена единица, обозначенная физиологическим эквивалентом рентгена (ФЭР).
Из видов ионизирующих излучений наиболее важное практическое значение имеют, помимо первичных космических лучей, рентгеновы лучи, α-, β- и γ-лучи, быстрые и медленные нейтроны. Жесткие рентгеновы лучи, у-лучи и нейтроны глубоко проникают в ткани животного организма и обозначаются, как проникающая радиация. Возникновение лучевой болезни в основном обусловлено действием этой группы лучей.
В мирное время наибольший интерес приобретает действие малых доз ионизирующих излучений. Они могут при одних условиях явиться стимулирующими, а при других — повреждающими агентами. По мнению Е. И. Бакина, находящиеся в животном организме естественные излучатели (40К, 14С, 3Н и др.) оказывают стимулирующее влияние на течение жизненных функций во время раннего периода развития организма, в последующие годы они повреждают клетки и ткани. Образно говоря, это лучи жизни, старения и смерти. Все виды излучений действуют на биосубстрат, вызывая процесс ионизации. Действующим началом их являются первичные и вторичные электроны, протоны и а-ча-стицы. Все они имеют общие черты сходства физиологического действия.
При радиационных поражениях имеет значение не только доза и мощность облучения, но также видовая, возрастная, половая и тканевая чувствительность организма к ионизирующим излучениям.
По наблюдениям, проведенным в лаборатории И. Т. Курцина на собаках с условными рефлексами по Павлову и хроническими фистулами внутренних органов, течение и исход лучевой болезни в значительной степени определяются, во-первых, типологическими свойствами высшей нервной деятельности и, во-вторых, исходным функциональным состоянием высших отделов центральной нервной системы организма перед облучением: чаще погибают собаки слабого и сильного неуравновешенного типа высшей нервной деятельности, а также когда облучение производится на фоне ослабления коры больших полушарий у собак с экспериментально вызванным неврозом.
При лучевой болезни в организме пет таких клеток И тканей, органов и систем органов, которые бы не подвергались радиационному повреждению. Повреждаются и органы пищеварения. Подробно этот вопрос изучен на собаках с хроническими фистулами, при общем тотальном облучении. Животные облучались от двух сдвоенных рентгеновских аппаратов, доза во всех случаях равнялась 350 рентгенам. Развитие лучевого поражения определялось по картине крови и ряду других показателей, в том числе по состоянию высшей нервной деятельности.
Опыты показали, что при таком виде и таких условиях облучения изменяется деятельность всех слюнных желез. Однако в динамике изменений секреторного процесса на действие пищевых и отвергаемых веществ со стороны разных желез отмечается некоторая особенность, обусловленная, в частности, степенью радиационного поражения. Наиболее характерной формой расстройств деятельности околоушных желез при легкой степени лучевой болезни является гиперсекреторная. При действии пищевых веществ гиперсекреторная реакция железы сохраняется в течение нескольких месяцев, тогда как при действии отвергаемых веществ (кислота) — не более одного месяца после облучения. При тяжелой форме лучевого поражения характерно резкое ослабление секреторной деятельности слюнных желез, причем главным образом изменения подъязычных и подчелюстных желез обусловливаются радиационным повреждением рефлекторных механизмов регуляции как условнорефлекторных, так и врожденных, и касаются они как количественной стороны секреции, так и ферментативных свойств слюны (появление амилазы).
При внутреннем облучении и вызываемом внутривенным введением радиоактивных веществ (фосфора, кобальта, стронция и др.) в зависимости от дозы наблюдается то возбуждающий, то тормозящий секреторный эффект на условно- и безусловнорефлекторные раздражения.
Итак, после внешнего или внутреннего, тотального или локального облучения организма, если доза и мощность ионизирующей радиации превышают предельно допустимые величины, возникают длительные функциональные расстройства слюнных желез. Механизм их обусловлен как прямым действием ионизирующего излучения на нервно-секреторный и сосудистый аппарат желез, так и опосредованным через нервную и гуморальную системы регуляции. Значительным изменениям подвергается условнорефлекторпый механизм регуляции деятельности слюнных желез. Изменения обнаруживаются уже в первые 10—15 с действия ионизирующего фактора: когда собаки при тотальном облучении получали всего лишь около двух рентген, развитие их было волнообразным: в латентном периоде лучевой болезни и в период выздоровления условнорефлекторное слюноотделение бывает повышенным, а в начале болезни и особенно в разгаре ее преобладают явления торможения условных слюноотделительных рефлексов.
Современные знания о секреторных расстройствах желудка при радиационных поражениях организма получены при клинических наблюдениях на людях, собраны в экспериментах на животных.
Клинический материал, несмотря на пестроту результатов и трудность сопоставления, позволяет выделить следующие три принципиальных положения: 1) ионизирующая радиация, на какую бы часть организма она ни действовала (желудок, печень, шея и др.), во всех случаях изменяет секреторный процесс желудка; 2) в малых дозах она стимулирует деятельность желудочных желез при пониженной активности и угнетает деятельность желез при гиперфункции; 3) в механизме функциональных изменений желудочных желез под влиянием ионизирующей радиации принимает участие нервная система, прежде всего вегетативный отдел ее, а также железы внутренней секреции.
У лиц, имевших длительный контакт с малыми дозами радиации, были также установлены нарушения секреторной функции желудка преимущественно в сторону ее угнетения.
Подтверждая эти данные в опытах на животных, экспериментальная радиационная физиология и радиационная патология установили и ряд других положений, важных в теоретическом и практическом отношениях. Так, после облучения у собак живота развивается лучевая болезнь, с первого дня которой вплоть до гибели животного деятельность желудочных желез резко угнетается в сложнорефлекторной фазе секреции, снижается переваривающая активность сока, степень его кислотности (Ю. Н. Успенский). При развитии лучевой болезни расстройства желудочной секреции имеют волнообразный характер количественных и качественных (пепсин, кислота) свойств, удлинение латентного периода возбуждения желез, изменение типа кривой секреции и продолжительности секреторного процесса на пищевое раздражение, появление спонтанной секреции, зависимость степени секреторных расстройств от тяжести лучевой болезни и наличия параллельно развивающихся нарушений высшей нервной деятельности. Выраженность секреторных расстройств в зоне малой кривизны бывает более яркой, чем в зоне дна и тела желудка. Аналогичные изменения отмечаются как после тотального облучения, так и после облучения живота.
Значительные изменения секреторной функции желудочных желез возникают и при внутреннем облучении— поступлении в организм радиоактивных веществ через желудочно-кишечный тракт или дыхательный аппарат. Токсическое поражение, вызываемое продуктами ядерного деления урана, вызывает, в зависимости от дозы, острую или хроническую форму лучевой болезни, и соответственно этому различные (гипо- и гиперсекреторные) расстройства желудочной секреции. То же самое возникает и при отравлении радоном, радиоактивным полонием. В механизме секреторных расстройств, возникающих при внутреннем облучении и интоксикации, должны учитываться помимо нарушений механизма рефлекторной и гуморальной регуляции деятельности желез также и нарушения, связанные с непосредственным токсическим влиянием веществ на секреторные клетки. Характер и продолжительность радиационных расстройств желудочной секреции находятся в зависимости от многих факторов, особенно от дозы ионизирующей радиации, типа нервной системы и функционального состояния ее высших отделов в момент облучения.
При функциональном ослаблении коры мозга нарушения деятельности желудочных желез, как и течение лучевой болезни, бывают ярко выраженными.
В литературе имеются факты, доказывающие большую чувствительность поджелудочной железы к ионизирующим излучениям. Уже с первых дней после облучения и развития острой лучевой болезни микроскопически в поджелудочной железе животных обнаруживаются структурные изменения, уплотнения и мелкие очаги жировой дистрофии с небольшой лимфацитарной инфильтрацией и другие дистрофические явления. Возникают количественные и качественные, главным образом ферментативные изменения секреции. Так же как и желудочная, поджелудочная секреция изменяется волнообразно, гиперфункциональная форма по ходу развития лучевой болезни сменяется гипофункциональной, и наоборот. Возникающие изменения секреции обусловлены главным образом расстройством нервно-гуморального механизма регуляции. Установлена зависимость этих изменений от радиационных форм повреждений высшей нервной деятельности, преобладания тормозных или раздражительных процессов в коре мозга. Словом, под влиянием ионизирующих излучений происходит резкое и длительное нарушение внешнесекреторной функции поджелудочной железы, которое проявляется в гипофункциональной, гиперфункциональной или смешанной форме, неустойчивости секреторной реакции на один и тот же пищевой раздражитель, волнообразности колебаний уровня секреции, изменении соотношения между количеством и качеством секрета, отделяющегося на различные сорта пищи, извращении типа секреторных кривых, возникновении инертности хода секреторного процесса, удлинении латентного периода сокоотделения, изменении соотношения активности ферментов на один и тот же пищевой раздражитель, нарушении относительного параллелизма отделения ферментов (явление диссоциации), возникновении спонтанной секреции.
Экспериментальные данные о биохимических изменениях, возникающих в печени под влиянием ионизирующей радиации и ведущих к нарушениям процессов в печени и во всем организме, давали основание предположить, что при радиационных поражениях должны быть серьезные расстройства со стороны желчеобразо-вательной и желчевыделительной функций. Предположение подтвердилось. В желчи собак после внешнего облучения рентгеновыми лучами в дозе 250 рентген концентрация холатов и билирубина в первые дни повышается, в разгаре лучевой болезни резко снижается и вновь повышается до исходного уровня, а иногда и выше в период выздоровления. Волнообразно изменяется концентрация желчных кислот. При локальном облучении печени малыми дозами секреция желчи увеличивается, а большими — уменьшается. Неустойчивый волнообразный характер спонтанной секреции желчи отмечается у собак, пораженных радиоактивным полонием. Наряду с изменением желчеобразования наступают также нарушения эвакуации желчи в кишку. Они характеризуются волнообразностыо, что связано с чередованием гипер- и гипофункциональных форм расстройств желчевыделительного аппарата, в частности изменениями моторной активности желчного пузыря и сфинктера Одди. Если облучение собак произвести на фоне функционально ослабленной коры больших полушарий (на фоне невроза), расстройства желчеобразо-вательной функции печени бывают более глубокими и продолжительными.
Наиболее обстоятельную характеристику нарушений секреторного процесса кишечных желез, возникающих под влиянием ионизирующих излучений, дали результаты хронических опытов, проведенных на собаках с изолированными петлями кишечника по методам Тири, Тири-Велла и Бабкина.
Лучевая болезнь, вызываемая облучением живота собак 350—660 рентген, сначала повышала, а затем понижала секрецию сока, что создавало на протяжении нескольких недель развития заболевания волнообразный характер в динамике секреторных расстройств (Ю. Н. Успенский). Изменения количества сока сопровождались увеличением амилолитических свойств секрета. Отмечались волнообразные изменения ферментативных свойств кишечного сока (энтерокиназы, фосфатазы, сахаразы, липазы и пептидаз) у облученных собак (М. Ф. Нестерин, А. П. Попов).
Волнообразный характер секреторных расстройств, зависимость степени нарушений секреторного процесса от дозы и мощности ионизирующего излучения, появление обильной спонтанной секреции, диссоциация в ферментообразовании и другие явления, возникающие после внешнего, тотального или локального облучения, имеют место и при внутреннем облучении и отравлении организма полонием, продуктами ядерного деления урана (С. Р. Перепелкин).
Одновременный анализ функционального состояния кишечных желез и высшей нервной деятельности показал, что секреторные расстройства кишечника могут быть результатом непосредственного радиационного поражения самих желез и нервно-сосудистого аппарата кишечника и как следствие нарушений механизма нервно-гуморальной регуляции секреторной функции кишечника, особенно со стороны высших мозговых центров.
Так, при лучевой болезни, вызванной облучением на фоне невроза, нарушения процесса секреции бывают более глубокими и продолжительными, чем при лучевой болезни, вызванной облучением на фоне нормального функционального состояния высших отделов центральной нервной системы.
Среди изменений работы пищеварительного аппарата при внешнем и внутреннем радиационном поражении организма важное место занимают нарушения процесса всасывания в тонком кишечнике. В основном оно замедляется, но часто замедление сменяется усилением, что придает динамике расстройств всасывательной функции кишечника волнообразный характер. Эти изменения могут быть обусловлены, с одной стороны, различной степенью радиационных повреждений местных биохимических и физико-химических процессов в ворсинках кишечника, в частности процессов окислительного фосфорилирования и мембранного пищеварения, с другой — нервно-гуморального механизма регуляции процесса всасывания. Существенное значение имеет при этом его регуляция высшими отделами центральной нервной системы. Наиболее значительные и длительные изменения процесса всасывания бывают при функциональном ос лаблении коры больших полушарии головного мозга и невротических нарушениях и нарушением в связи с этим корково-подкорковых взаимоотношений.
При лучевой болезни резко усиливается экскреторная функция желудочно-кишечного тракта, главным образом за счет увеличения выделения продуктов расщепления белковых молекул.
Боль и ее влияние на пищеварение
В конце XVIII в. Э. Дарвин писал, что неприятные эмоции и боль оказывают угнетающее влияние на процессы пищеварения и усвоения пищи. С. М. Дионесов, исходя из процесса видового приспособления организмов к условиям существования, полагает, что под влиянием боли должна была выработаться способность к задержке пищеварительных процессов. Активность пищеварительной системы в это время непроизводительно отвлекала бы излишние массы крови от органов, принимающих участие в осуществлении защитно-оборонительных реакций.
Проведенными исследованиями установлено, что на болевое раздражение реагируют слюнные железы. Так, О. В. Овсянников и С. И. Чирьев при болевом раздражении индукционным током центрального конца перерезанного седалищного нерва наблюдали у собаки обильное слюноотделение из подъязычной и подчелюстной желез, причем слюна была жидкой. Другие исследователи также наблюдали повышенное слюноотделение при болевом раздражении, наносимом па кожу.
А. А. Дудорова в опытах на собаках показала, что слюнные железы, находящиеся в состоянии относительного покоя, при растяжении баллоном ампулы прямой кишки начинают отделять слюну. Это указывает на то, что раздражение интерорецепторов (барорецепторов) прямой кишки переводит их в активное состояние. Необходимо при этом отметить, что на растяжение стенки прямой кишки реагируют преимущественно смешанные железы. Что же касается околоушных желез, то они либо вовсе не отвечают, либо отвечают слабо. Выявилась зависимость интенсивности слюноотделения от степени растяжения стенки прямой кишки и при этом было установлено, что после прекращения действия раздражителя выделение слюны прекращается. Химические исследования показали, что выделяемая на растяжение ампулы прямой кишки слюна содержит мало плотных веществ. Иной результат дали опыты с растяжением анальных сфинктеров (экстероцептивное раздражение). Выяснилось, что даже сильное растяжение их не сопровождалось сколько-нибудь заметным слюноотделением. Вместе с тем у тех же животных растяжение ампулы прямой кишки (интероцептивное раздражение) вызывало интенсивное слюноотделение. Раздражение электрическим током кожи вызывало незначительное слюноотделение, т. е. реакция была сходной с растяжением анальных сфинктеров.
Слюнные железы реагируют спонтанным слюноотделением не только на растяжение ампулы прямой кишки, но и мочевого пузыря. С прекращением растягивания слюноотделение постепенно снижалось и затем вовсе прекращалось. По содержанию хлоридов, плотного остатка слюна, отделяемая на растяжение стенки мочевого пузыря, напоминала слюну, отделяемую на пилокарпин.
С. А. Фроловым было показано, что в условиях хронического опыта раздражение индукционным током желудка и кишок со стороны серозной оболочки у собак с вживленными электродами переводило смешанные железы из состояния покоя в деятельное состояние. Раздражение париетальной плевры током сопровождалось незначительным слюноотделением.
Те же воздействия на экстеро- и интерорецепторы, ко применявшиеся на фоне активной деятельности слюнных желез, обусловленной введением пилокарпина, вызывали снижение интенсивности слюноотделения. Однако эффект зависел от того, каким был исходный уровень саливации при нанесении раздражения: при низком уровне наблюдалось усиление слюноотделения, при более высоком — отчетливо проявлялось его торможение. Следовательно, одно и то же воздействие, наносимое на фоне одного исходного состояния слюнных желез, вызывает возбуждение (стимуляция), на фоне другого — торможение.
Произведенный анализ экстеро- п интероцептивных влияний на деятельность слюнных желез, проведенный различными способами (фармакоанализ, денервация), позволил установить, что в передаче нервных импульсов принимают участие разные отделы центральной нервной системы и в том числе кора головного мозга. Эфферентные импульсы из центральной нервной системы направляются к слюнным железам по парасимпатическим и симпатическим нервам.
И. П. Павлов наблюдал торможение секреции слюны у собаки при раздражении брюшных внутренностей.
А. Нечаев у собак с басовской фистулой наблюдал при сильном раздражении седалищного или большебер-цового нервов угнетение сокоотделения в желудке. Ман-телли отметил у человека с фистулой желудка, что болевое раздражение тормозило уже имевшуюся секрецию и препятствовало ее началу, если желудок находился в покое.
С. М. Дионесов, обобщая накопленный материал исследований, выдвинул и обосновал следующую схему угнетения секреторной деятельности желудка при болевых воздействиях. Сразу же вслед за нанесением животному болевого раздражения из центральной нервной системы к различным органам, в том числе и желудку, направляется поток импульсов, изменяющих функциональное состояние этих органов. Помимо непосредственного рефлекторного влияния нервной системы па секреторный аппарат желудка деятельность последнего изменяется и гуморальным путем. При действии болевого раздражителя происходит выделение специфических и неспецифических продуктов, способных гуморалыю воздействовать на деятельность желез желудка и вызывать снижение их активности. К числу таких продуктов следует отнести некоторые гормоны, в частности инсулин, адреналин и питуитрин.
Показано, что воздействие электрическим током на участок костальной плевры вызывает торможение желудочной секреции, возбуждаемой как мнимым кормлением мясом, так и дачей 200 г мяса. Однако в опытах, в которых болевое раздражение наносилось до кормления животного, желудочные железы переходили из состояния покоя в деятельное состояние.
М. Я. Гукова наблюдала у больных с заболеваниями внутренних органов во время приступов сильных болей угнетение секреции желудочного сока.
С. С. Полтырев выявил у собак значительное торможение желудочной секреции при нанесении на стенку мочевого пузыря индукционного тока, при этом торможение было пропорционально силе тока. После прекращения раздражения сокоотделение устанавливалось на исходном уровне. Необходимо отметить, что раздражение током мочевого пузыря вызывало спастическое сокращение его стенки, чем и был обусловлен болевой синдром.
Применив метод хромоскопии, было установлено, что болевое раздражение стенки прямой кишки, мочевого пузыря угнетает не только сокоотделение, но и задерживает выделение краски нейтральрот железами желудка. Кроме того, в отделяемом желудочном соке возрастало содержание мочевины, креатинина и молочной кислоты (Е. А. Бродская).
Вопрос о влиянии болевого раздражения на поджелудочную секрецию изучен недостаточно. Однако известно, что болевое раздражение угнетает сокоотделение, возбуждаемое как соляной кислотой, вводимой в тонкую кишку, так и секретином, вводимым внутривенно. С. С. Серебреников считает, что торможение распространяется главным образом на нервную фазу, не затрагивая гуморальной фазы поджелудочной секреции.
Болевое раздражение оказывает влияние на желчеобразование и желчевыделение. В опытах с раздражением электрическим током чувствительных нервов (седалищного и др.), желчеотделение стимулировалось или угнеталось в зависимости от интенсивности воздействия. Торможение желчеотделения было отмечено и при электрическом раздражении кожи.
Установлено, что при раздражении кишечной стенки электрическим током в условиях хронического опыта наступало значительное снижение желчеотделения в период наивысшей пищевой секреции; на фоне низкой пищевой секреции тормозной эффект был менее выражен. При нанесении болевого раздражения в период накопления желчи в желчном пузыре, выведение ее в двенадцатиперстную кишку задерживается; в опытах, в которых раздражение приурочивалось к периоду усиления выхода желчи, наблюдалось либо усиление этого процесса, либо незначительное его ослабление. На основании этого можно сделать заключение, что сфинктер Одди, находясь в расслабленном состоянии, реагирует га болевое воздействие слабо, поэтому усиливающееся в это время выделение желчи обусловлено активными сокращениями желчного пузыря.
На собаках с фистулой по методу Тири — Велла было установлено стимулирующее влияние болевого раздражения кожи током на секрецию кишечных желез, причем стимуляция появлялась, когда раздражение предшествовало орошению петли кишки 0,2%-ным раствором соляной кислоты, и в случае, когда оно наносилось сразу после орошения или на фоне уже развившейся секреции (С. С. Серебреников). Вместе с тем некоторые авторы наблюдали не усиление кишечной секреции, а торможение ее вплоть до полного прекращения сокоотделения при нанесении болевого раздражения на кожу. Больше того, угнетение секреции отмечалось не только в период болевого раздражения, но и после прекращения его в течение некоторого времени.
Нанесение болевого раздражения собакам с денер-вированной петлей тощей кишки в период так называемой паралитической секреции кишечного сока изменяет его количество по-разному: в первые 1,5 месяца после денервации кишечная секреция претерпевала некоторые изменения, а спустя 2,5 месяца изменения отсутствовали. Однако через 6 месяцев, по мере регенерации нерв-пых волокон, кишечные железы на болевое раздражение отвечали торможением, т. е. так же, как и у собак с интактной петлей кишки. При проведении опытов было отмечено, что возможно образование у собак услов-норефлекторной болевой реакции со стороны кишечных желез. Итак, в реализации эффекта болевого раздражения на железы тонкой кишки принимает участие нервная система, включая кору головного мозга, гормоны надпочечников (адреналин) и гипофиза (питуитрин)
Угнетение кишечной секреции было выявлено у собак при нанесении болевого раздражения не только на кожу, но и на интерорецепторы тощей, слепой и ободочной кишок, причем торможение проявлялось на фоне возбуждения кишечной секреции как механическим воздействием, так и химическим (соляная кислота). Болевое воздействие снижало уровень кишечной секреции, возбуждаемой и пилокарпином.
Извращение периодической секреции кишечного сока у голодных собак отмечалось при кратковременном раздражении кожи электрическим током.
Болевое воздействие оказывает влияние не только на деятельность пищеварительных желез, но и на двигательную способность желудка. Еще в 1890 г. В. М. Бехтерев и Н. А. Миславский нашли, что болевое раздражение (укол иглой или электрическое раздражение) приводило к временной остановке или ослаблению ритмических сокращений привратниковой части и движений стенки желудка.
Специальными исследованиями было установлено значение исходного функционального состояния органа для проявления влияния болевого раздражения. В частности, в опытах, проведенных на японских макаках, при раздражении центрального конца седалищного нерва наблюдалось повышение тонуса пустого желудка, если до нанесения раздражения он был низок, и понижение тонуса, если он был высок.
При болевых раздражениях кожи происходит изменение эвакуаторной способности желудка, однако оно проявляется по-разному в зависимости от того, какое испытуемое вещество вводится в полость желудка. В частности, эвакуация воды замедляется, а соляной кислоты ускоряется. В осуществлении этих влияний участвует парасимпатическая нервная система. В механизме влияния двухчасового болевого раздражения на деятельность желудка выявлено значение блуждающих и чревных нервов. При частичном повреждении некоторых подкорковых структур (головки хвостатых ядер, задних вентро-латеральных ядер зрительных бугров) характер реакции желудка на продолжительное болевое раздражение кожи изменяется. В реакции на болевое раздражение кожи участвует и кора головного мозга. Болевое раздражение вызывает у собак уменьшение амплитуды исходного |3-ритма электроэнцефалограммы и явление синхронизации в сенсорно-моторной, теменной и затылочной областях коры головного мозга. Повторные продолжительные болевые раздражения сопровождаются условнорефлекторными изменениями желудочной секреции на мясо в виде уменьшения количества сока в первые два часа и снижением его кислотности.
Выяснено, что в развитии торможения желудочной секреции при длительном болевом раздражении кожи участвуют глюкокортикоиды.
Отмечено, что при болевом раздражении интероре-цепторов у собак происходит замедление эвакуации из желудка в двенадцатиперстную кишку 0,5%-ного раствора соляной кислоты, 3%-ного раствора крахмала, что обусловлено как понижением тонуса мускулатуры желудка, так и повышением тонуса пилорического сфинктера.
Болевые раздражения изменяют тонус гладкой мускулатуры и тонкой кишки, что проявляется в большинстве случаев ослаблением маятникообразных движений, прекращением ритмических и перистальтических движений кишки, снижением ее тонуса. Однако в отдельных опытах удавалось наблюдать и противоположный эффект, что, по-видимому, зависело от различного исходного состояния мышечного аппарата кишки в момент болевого воздействия. В механизме влияния болевого раздражения важная роль принадлежит чревным нервам, так как после перерезки их болевое раздражение уже не меняет характера двигательной активности тонкой кишки.
В 1904 г. В. Н. Болдырев отметил при болевом раздражении нарушение периодической двигательной деятельности тонкой кишки. Это наблюдение получило подтверждение и в опытах С. И. Франкштейна. Болевое раздражение кожи электрическим током сразу же обрывало голодные движения кишки. Вместе с тем было отмечено, что при нанесении болевого раздражения в дистальной части кишечника наблюдается усиление двигательной активности при одновременном ослаблении тонуса запирательных мышц, в результате чего при болевом раздражении содержимое дисталь-ного отдела может быть внезапно выброшено наружу. Это биологически целесообразно для организма, мобилизующегося для отпора разрушительному фактору.
Исследованиями Р. О. Файтельберга с сотрудниками показано, что под влиянием болевого раздражения происходит ускорение всасывания глюкозы в желудке и тонкой кишке. Ускорение всасывания происходит и условнорефлекторным путем — на треск индуктория. Это свидетельствует об участии коры головного мозга в осуществлении влияния боли на скорость всасывания в желудке и кишечнике. Опыты показали, что в механизме влияния боли на процесс всасывания определенная роль принадлежит и эндокринным факторам, в частности адреналину, тироксину и питуитрину. На процесс всасывания в тонкой кишке оказывают влияние вагус и чревный нерв. При выключении их реакция всасывательного аппарата на действие болевого раздражения изменяется.
Итак, боль вызывает существенную перестройку в работе органов пищеварения, причем она может стать причиной значительных пищеварительных расстройств. Стойко удерживающаяся у человека боль приводит к угнетению деятельности пищеварительных желез, двигательного аппарата желудочно-кишечного тракта, а также резорбтивных процессов в желудке и кишечнике.
Глава 13
ВОЗРАСТНЫЕ ОСОБЕННОСТИ ПИЩЕВАРЕНИЯ И ПИТАНИЯ
Созданное учение о работе органов пищеварения способствует успешному изучению многих вопросов возрастной физиологии пищеварения, крайне важных для разработки проблем рационального питания детей и профилактики желудочно-кишечных заболеваний.
Пищеварительный аппарат к моменту рождения сформирован в структурно-функциональном отношении не полностью. С возрастом происходит созревание структуры и функций желудочно-кишечного тракта и его деятельность становится более совершенной. Заканчивается процесс формирования к зрелому возрасту. Исходя из этого питание детей должно соответствовать функционально-структурным особенностям пищеварительной системы в разные возрастные периоды. Если же такого соответствия не будет, то могут возникать пищеварительные расстройства с последующим нарушением обменных процессов.
В педиатрии различают три периода детства: первый— до 2,5 лет, второй — от 2,5 до 7 лет и третий — от 7 до 14 лет.
Период молочного вскармливания — самый ранний. Следует отметить, что в литературе по вопросу о характере функционирования различных пищеварительных желез в этом периоде имеются противоречивые данные. Однако большинство считает, что в период новорожденности слюнные железы выделяют незначительное количество слюны и при этом она не содержит муцина или бедна им. В этот период в слюне удается обнаружить только один фермент — птиалин, а мальтаза отсутствует. Несмотря на незначительное выделение слюны, значение ее огромно. Под ее влиянием молоко, поступающее в желудок, створаживается мелкими и нежными хлопьями, что не безразлично для процесса желудочного пищеварения. С возрастом увеличивается выделение слюны и в ней появляются муцин и фермент мальтаза.
Слюноотделение усиливается в период прорезывания зубов. Это имеет большое приспособительное значение, так как с появлением постоянных зубов расширяется возможность потребления сухой и твердой пищи, что, естественно, требует большего количества слюны (А. А. Маркосян).
Существенные особенности имеются в работе желудка в первые месяцы жизни ребенка.
В течение первого года жизни основная часть желудка— дно составляет лишь четверть длины всего желудка, в то время как у взрослого она равняется одной трети. Желудочные железы у ребенка короче и шире, чем у взрослого; главные секреторные клетки небольшие и низкие; обкладочные клетки имеют неправильную форму и в значительной части лишены бокаловидных кист. В течение всего периода детства происходит непрерывное увеличение количества и размера секреторных клеток. У новорожденного насчитывается около 2 000 000 секреторных клеток, 10-летнего—17 000 000, 15-летнего— 22 500 000 и у взрослого человека их около 25 млн.
У детей первого периода детства и, в частности, первых месяцев жизни, сок, выделяемый желудочными железами, либо вовсе не содержит свободной соляной кислоты, либо количество ее невелико. В соке обнаружена молочная кислота (10—20 титр, ед.), пепсина мало, но он богат ферментом химозином, участвующим в створаживании казеина молока. Химозин проявляет свое действие в слабощелочной среде.
Несколько иные данные приводятся в клинической литературе. Так, Ю. Ф. Домбровская с соавторами (1970) указывает, что желудочное пищеварение у доношенного ребенка устанавливается тотчас же после рождения. У новорожденного желудочный сок содержит все составные части: ферменты (пепсин, химозин, липазу), молочную кислоту, соляную кислоту, поваренную соль. Состав желудочного сока у детей первого месяца жизни: общая кислотность— 3,8 титр, ед., свободная соляная кислота— 0,8—4,5 титр, ед., пепсин — 2—8 усл. ед., химозин—32 ед., липаза —4,2—10,2 ед. (М. С. Маслов). Однако и эти цифры свидетельствуют о низком содержании в желудочном соке свободной соляной кислоты и соответственно низкой активности пепсина. При питании женским молоком здоровые грудные дети выделяют желудочный сок с меньшей кислотностью и активностью ферментов, чем при вскармливании коровьим молоком (А. Ф. Тур).
У детей первого месяца жизни количество отделяемого сока невелико, что обусловлено видом и запахом пищи, обстановкой, связанной с приемом пищи, а также тем, что химические и механические раздражители в этом возрасте не оказывают выраженного стимулирующего влияния на деятельность желудочных желез.
В ранний период у детей наблюдается ряд особенностей в кишечном пищеварении. А. А. Маркосян указывает, что в это время поджелудочная железа практически не функционирует и сока не выделяет. Объясняется это тем, что в желудочном соке отсутствует свободная соляная кислота — стимулятор образования гормона секретина в тонкой кишке. В связи с отсутствием или недостаточным образованием секретина способность железы вырабатывать поджелудочный сок невелика.
В противоположность А. А. Маркосяну С. И. Гальперин отмечает, что у новорожденного поджелудочная железа выделяет много сока, и ее усиленная деятельность как бы восполняет недостаточную активность желудочных желез. Однако при этом нельзя не учитывать, что поджелудочный сок беден ферментами, в частности амилазой; что же касается липазы, то она содержится в форме зимогена.
М. С. Маслов считает, что недостаточная липолитиче-ская активность сока может компенсироваться наличием липазы в молоке матери. О протеолитической активности сока имеется мало данных. Домбровская с соавторами указывает, что у ребенка с рождения поджелудочный сок содержит трипсин, хотя активность его низкая.
Нерешенным остается вопрос, образуется ли в слизистой оболочке тонкой кишки гормон панкреозимин, стимулирующий образование в поджелудочной железе ферментов.
В последние годы выяснено, что кишечные железы выделяют сок, содержащий с первых недель жизни ребенка ферменты: энтерокиназу, щелочную фосфатазу, эрепсин, мальтазу, лактазу, нуклеазу (Л. С. Фомина). Образование фосфатазы с первых часов жизни ребенка подтверждает значение ее в качестве фактора, обеспечивающего всасывание в кишечнике продуктов гидролиза белков, углеводов и жира. У детей более старшего возраста в кишечном соке обнаруживается и липаза.
В раннем возрасте желчь бедна кислотами и в особенности гликохолевой, но богата слизью и пигментами. У новорожденных печень находится в состоянии функциональной недостаточности, однако с возрастом количество выделяемой желчи увеличивается и на 1 кг массы тела становится в 4 раза больше, чем у взрослого человека.
Всасывательная деятельность — главная функция тонкой кишки. В ней происходит всасывание продуктов гидролиза белка, жира, углеводов и отчасти солей. В толстых же кишках всасываются вода, железо, фосфор и щелочи. Здесь, как и у взрослых, в связи с наличием микрофлоры (кишечная палочка, энтерококк, дрожжевая флора) происходят гнилостные и бродильные процессы. Однако гниение выражено слабо. Ряд бактерий участвует в синтезе витамина К и витаминов группы В. Следует отметить, что уже в первые недели жизни ребенка происходят изменения в составе микрофлоры.
В период молочного вскармливания двигательная деятельность желудочно-кишечного тракта имеет некоторые особенности. В частности, кардиальный сфинктер слабо развит. Недоразвитие его при слабой мускулатуре желудка способствует частому срыгиванию. Повышение тонуса мускулатуры желудка у ребенка этого возраста может обусловливать пилороспазм. При грудном вскармливании желудок опорожняется через 2—3 ч, а при вскармливании коровьим молоком — через 3— 4 ч. Моторика кишечника у детей раннего возраста энергичная, но вместе с тем отличается непостоянством. Она резко усиливается под влиянием как местного раздражения, так п внешних воздействий (перегревание, резкое звуковое раздражение, крик, общее двигательное беспокойство и пр.). Весь путь по тонкому кишечнику пищевая кашица проходит за 12—30 ч при грудном вскармливании, а при искусственном продолжительность увеличивается.
В первые дни после рождения нормальная и естественная пища ребенка — молозиво и молоко матери. Молозиво отделяется в течение нескольких дней после родов, а затем переходит в молоко. Молозиво очень богато белком— 9—12%, в нем содержится 4—5% жира и 0,4—0,5% солей. Калорийность молозива в два раза выше, чем молока. Плотность его равна 1,050—1,060. В нем обычно содержатся ферменты (каталаза, пероксидаза), антитела и антитоксины, имеющие важное общебиологическое значение. Антитела, поступающие с молозивом в пищеварительный тракт новорожденного, а также глобулины, с которыми связано образование антител и антитоксинов, способны предохранять организм новорожденного от заражения и прежде всего кишечной палочкой. Вместе с тем доказано, что через молозиво к новорожденным переходят иммунизирующие вещества от активно или пассивно иммунизированных матерей.
После перехода молозива в молоко оно продолжает сохранять высокие питательные и в некоторой мере биологические свойства. При естественном вскармливании молоко здоровой матери является единственной естественной физиологической пищей ребенка в первые 4— 5 месяцев жизни. Однако в случае неправильного вскармливания, переедания у детей могут наступить пищеварительные расстройства, отрицательно сказывающиеся па обмене веществ, общем состоянии, темпе роста и развития. На успешность кормления до известной степени влияет режим кормящей матери, образ ее жизни и труда, характер питания. В частности, кормящая мать должна избегать употребления пахучих веществ (лук, чеснок придают молоку специфический вкус и запах, в связи с чем у ребенка может снизиться аппетит), спиртных напитков.
Ребенку в первую четверть года требуется (в среднем на 1 кг массы тела) 110—120 калорий, во вторую четверть—100—110 калорий, а в третью и четвертую четверти — 90—110 калорий.
Важное гигиеническое значение имеет регулярность кормления грудью.
Период смешанного вскармливания. Переход от молочного питания к смешанному вызывается необходимостью. Дело в том, что в течение первого года жизни ребенка масса тела быстро нарастает. Соответственно с каждым месяцем все более возрастает потребность организма в питательных веществах. На 5—6-м месяце жизни грудное молоко не может полностью удовлетворить потребность организма в питательных веществах. Из клинической практики известно, что затянувшееся свыше 6—7 месяцев кормление грудным молоком нередко ведет к развитию у детей анемии, запоров и снижению тургора тканей. Причиной анемии считают недостаточное содержание в грудном молоке железа и резкое снижение его запасов, ранее имевшихся в организме ребенка. Не случайно такую форму развивающейся анемии называют железодефицитной. Однако это не дает права снижать достоинства грудного молока для детей 5—6 месяцев и до года (табл. 23).
Таблица 23. Необходимые пищевые вещества и калорийность пищи для детей первого года жизни
Пищевые вещества,г/кг | ||||
| Вскармливание | белки | жиры | углеводы | Калорийностьккал |
| Естественное Смешанное Искусственное | 2,5 3,5 4,5 | 6,3 5,6 5,0 | 12,5 12,6 3,0 | 120 118 115 |
Грудное женское молоко содержит 17 аминокислот, необходимых для правильного развития ребенка. В состав жира входят ненасыщенные жирные кислоты (олеиновая кислота — 5,2%, линолевая—1,5%, стеариновая— 11,5%, пальмитиновая — 30%) и низшие ненасыщенные кислоты (5%)- В жире женского молока содержатся фосфатиды, в зрелом молоке 1,7%. Они (з основном лецитины) важны для ускорения всасывания жира в кишечнике, улучшения использования липидов в тканях и пр. В составе женского молока имеются многие макро- и микроэлементы: кальций, магнии калий, натрий, фосфор, хлор, сера, железо, кобальт, алюминий, цинк, марганец, йод, литий и др. Минеральные вещества входят в состав некоторых ферментов, ускоряют ферментативные процессы, участвуют в различных обменных процессах. В молоке содержатся различные витамины.
Для того чтобы избежать появления и развития анемии и других нарушений в организме ребенка, отрицательно влияющих на общее развитие и рост, его с 5—6-месячного возраста начинают прикармливать.
С этого момента на смену молочному приходит смешанное вскармливание.
Возникает вопрос — не следует ли переводить ребенка на смешанное вскармливание раньше, например с 2—3 месяцев? Дело в том, что в первые 3 месяца жизни пищеварительный тракт еще недостаточно сформирован и к тому же чувствительность его к необычным пищевым раздражителям очень высокая. С 5—6 месяцев прикармливание становится не только возможным, но и необходимым, поскольку к этому времени пищеварительная система претерпевает существенные преобразования. В частности, увеличивается емкость желудка с 30 мл при рождении до 100 мл к 3 месяцам и до 250 мл к году. Более интенсивно происходит слюноотделение, нарастает секреторная активность желез желудка, сокоотделение в желудке начинает протекать в обе фазы — сложнорефлекторную и нервно-химическую. В желудочном соке повышается содержание соляной кислоты при одновременном снижении концентрации молочной кислоты. С нарастанием кислотности желудочного сока создаются благоприятные условия для повышения активности пепсина как протеолити-ческого фермента. Активируется и кишечное пищеварение, причем преимущественно за счет усиления секреторной деятельности поджелудочной железы; выделяемый сю поджелудочный сок с его разнообразными ферментами способствует лучшему перевариванию всеч основных составных частей пищи в кишечнике. Все это создает необходимые предпосылки для того, чтобы с 5—6-месячного возраста начать прикармливать ребенка, проявляя при этом большую осторожность и осмотрительность.
Прикорм, каким бы он ни был, по своим вкусовым и другим качествам значительно отличается от грудного молока. Поэтому новую пищу начинают давать с небольших порций (30—50 г), постепенно увеличивая ее количество. Прикорм дается взамен одного кормления грудью. Это предупреждает, во-первых, от переедания, а во-вторых, в связи с повышением пищевой возбудимости ребенок охотнее принимает необычную пищу. Через 3—4 недели вводится второй прикорм, а в дальнейшем третий, четвертый и т. д. Так к 11 —12 месяцам ребенок полностью отлучается от груди. При таком постепенном отлучении у детей не возникают расстройства стула. Нередким затруднением при прикармливании является отказ ребенка от непривычной ему пищи. В таких случаях требуется терпение: иногда достаточно дать ребенку поголодать, чтобы через 3— 4 ч опять предложить ему ту же пищу, и он отнесется к ней совсем иначе.
В период молочного вскармливания единственным пищевым возбудителем является грудное молоко, а при искусственном вскармливании — коровье. Следовательно, в период молочного вскармливания вкусовой аппарат ребенка адаптирован только к молоку. Однако для того чтобы прикорм стал естественным пищевым раздражителем в период смешанного вскармливания, требуется физиологическая перестройка не только периферического конца вкусового анализатора, но и центрального. Вместе с тем из физиологии известно, что процесс приспособления к новым пищевым раздражителям осуществляется сравнительно медленно и при условии, что они повторяются многократно.
В том случае, если переход с молочного вскармливания к смешанному, с включением различных прикормов, протекал гладко и за этот период значительно расширились приспособительные возможности вкусового аппарата ребенка, можно ожидать, что в возрасте одного года он успешно справится с приемом более разнообразной и грубой пищи.
Питание детей от 1 года до 3 лет
При определении режима питания детей раннего возраста исходят из особенностей возраста. На этом этапе развития дети имеют более совершенную пищеварительную систему по сравнению с детьми грудного возраста. Увеличивается ферментативная активность пищеварительных соков, вкусовые восприятия становятся более дифференцированными, повышается приспособляемость организма. Постепенно совершенствуется и жевательный аппарат. В конце первого года ребенок имеет 6—8 зубов и может не только глотать, но достаточно хорошо пережевывать пищу. Все это позволяет разнообразить пищу и приблизить ее по составу, вкусу и консистенции к пище взрослых.

Для обеспечения полноценного питания необходимо соблюдать следующие принципы. Прежде всего пища, которую получает ребенок, должна отвечать его калорийным потребностям, быть качественной И полноценной, т. е. содержать необходимое количество белков, жиров и углеводов, минеральных солей и витаминов (табл.24). Белок —основной строительный материал, из которого строятся новые клетки организма. Они входят в состав антител, ферментов и гормонов, которые обеспечивают защитные силы организма и регулируют обмен веществ. Недостаточное поступление с пищей белков в этом возрасте неблагоприятно отражается на росте и развитии ребенка. Уместно отметить, что отрицательное влияние на функции нервной системы, печени, почек оказывает как недостаток, так и избыток белка. Задержка роста и развития наблюдается при недостатке ненасыщенных жирных кислот. При недостатке жиров возможно снижение иммунобиологической реактивности, а при избытке их нарушение обмена веществ, расстройство деятельности органов пищеварения. Необходимо считаться со вкусом и индивидуальными особенностями ребенка. Следует исключать из рациона плохо переносимую пищу, чтобы предупредить тяжелые реакции и, В первую очередь, аллергического характера.
При составлении рациона включают продукты, которые являются возбудителями деятельности пищеварительного аппарата. К их числу можно отнести молочнокислые продукты (кефир, простокваша, ацидофилин и др.); соки и пюре из ягод и овощей, муссы. В связи с нарастанием активности протеолитических ферментов желудочного сока, повышением его кислотности с 1,5 лет в ассортимент пищевых продуктов вводят ржаной хлеб и макаронные изделия.
В связи с продолжающимся развитием высшей нервной деятельности, с расширяющимися возможностями для образования временных связей (условных рефлексов) у детей в этот период развития важное значение приобретает режим питания и, в частности, регулярный прием пищи. И. П. Павлов придавал этому большое значение. Он считал необходимым создавать спокойную, приятную обстановку, направляя все внимание ребенка на еду. Ни в коем случае не может быть оправданным отвлечение ребенка разговорами и посторонними предметами.
Дети, воспитывающиеся в яслях, имеют преимущество перед детьми, воспитывающимися в домашних условиях. Они находятся на нормированном питании, осуществляемом под контролем медицинских работников, а также строго соблюдается режим ребенка. Кроме того, ребенок с пониженным аппетитом, питаясь в коллективе, быстрее проявит стремление не отставать от сверстников.
Питание детей дошкольного возраста. Исследованиями установлено, что у детей до шестилетнего возраста содержание соляной кислоты в желудочном соке еще не достигает уровня, характерного для детей школьного возраста, и колеблется в пределах 10—20 титр. ед. Содержание протеолитических ферментов сохраняется на том же уровне, как и у детей от 1 года до 3 лет. Двигательная активность желудка и кишечника осуществляется менее интенсивно, чем у детей раннего возраста. Емкость желудка к четырем годам достигает 600 мл, а с четырех до семи лет увеличение объема замедляется. В целом пищеварение и процессы всасывания соответствуют уровню, характерному для детей младшего школьного возраста (А. А. Маркосян).
С учетом отмеченных особенностей в работе органов пищеварения питание детей дошкольного возраста отличается от питания детей ясельного возраста (табл. 25).
В связи с быстрым развитием высшей нервной деятельности детей дошкольного возраста пища должна быть не только хорошо приготовленной и вкусной, но и красиво поданной. Это, безусловно, способствует повышению аппетита. При еде ребенок должен ощущать вкус пищи, и поэтому еда должна быть неторопливой. Торопливая еда не дает вкусовых ощущений и наслаждений, так как принятая пища очень непродолжительное время находится в полости рта.
Питание детей школьного возраста. У детей школьного возраста, особенно среднего и старшего, строение и деятельность пищеварительных органов мало отличается от таковых у взрослых. Поэтому при нормировании питания приходится учитывать в основном лишь следующие важные физиологические особенности: продолжающийся рост и развитие, увеличение массы тела, интенсивное развитие психической деятельности и мышечной массы, условия жизни и учебных занятий школьника.
Рациональное питание детей школьного возраста способствует нормальному физическому и умственному развитию и снижает утомляемость (табл. 26, 27, 28). Специально проведенными исследованиями установлено, что школьнику недостаточно утреннего домашнего завтрака, поэтому в школах нашей страны питание организовано так, что учащиеся получают горячие завтраки, не допуская большого перерыва между утренним домашним завтраком и обедом.
Для детей школьного возраста имеет большое значение соблюдение режима питания. Если пища принимается ежедневно в одни и те же часы, то организм привыкает (формируется стереотип) к таким приемам пищи и как бы подготавливается к ним, что способствует лучшему пищеварению. При беспорядочном питании пищеварение протекает хуже, так как нарушается деятельность коры больших полушарий головного мозга и вместе с этим слаженная работа пищеварительных органов. Кроме того, снижается аппетит. К снижению аппетита ведет и однообразная пища. Все это говорит о необходимости создания таких условий питания детям, которые всемерно способствовали бы их нормальному росту и развитию и предупреждали появление расстройств питания.и пищеварения.
Таблица 25. Средний суточный набор пищевых продуктов для детей от 5 до 6 лет
Наименованиепродуктов | Количество, г | Пищевыевещества, г | Калорийность, ккал | ||
белки | жиры | углеводы | |||
Молоко | 750 | 21 | 26,2 | 33,7 | 468,6 |
Творог | 50 | 6 | 4,25 | 1,65 | 70,5 |
Сметана | 10 | 0,21 | 2,85 | 0,31 | 26,2 |
Сир | 10 | 1,87 | 1,71 | 0,18 | 24,3 |
Яйцо | 25 | 2,25 | 2,42 | 0,07 | 31,8 |
Мясои мясопродукты | 105 | 13,86 | 2,73 | — | 79,3 |
Рыба и рыбопродукты | 50 | 5,85 | 0,15 | — | 25,37 |
Хлебпшеничный | 100 | 7,62 | 1,2 | 49 | 213,2 |
» ржаной | 40 | 2 | 0,4 | 17 | 81,6 |
Мука пшеничная | 15 | 1,4 | 0,15 | 11,45 | 54 |
» картофельная | 8 | 0,06 | — | 6,48 | 26,7 |
Крупа гречневая | 7 | 0,7 | 0,16 | 4,41 | 22,4 |
» манная | 5 | 0,48 | 0,03 | 3,5 | 16,65 |
» рисовая | 8 | 0,5 | 0,07 | 5,7 | 26,08 |
» геркулес | 5 | 0,54 | 0,3 | 3,05 | 17,5 |
Макароны | 10 | 0,93 | 0,08 | 7,09 | 33,6 |
Сладости | 10 | 0,5 | 1 | 6,7 | 38,8 |
Сахар | 60 | — | — | 57,3 | 234 |
Картофель | 250 | 3 | — | 35 | 155,8 |
Овощи разные | 300 | 3,12 | — | 13,2 | 66,9 |
Фруктысвежие | 200 | 0,4 | — | 20,02 | 84,46 |
» сухие | 10 | 0,2 | — | 10,01 | 42,23 |
Маслосливочное | 23 | 0,07 | 18,05 | 0,11 | 168,82 |
» растительное | 10 | — | 9,38 | — | 87,2 |
Поваренная соль | .8 | — | — | — | — |
Чай | 0,2 | — | — | ||
Кофе | 3 | — | — | — | — |
Какао | 0,5 | 0,1 | 0,09 | 0,19 | 2 |
Всего: | 72,65 | 71,22 | 286,12 | 2128 | |
Изних продукты: | |||||
молочные | 29,15 | 53,06 | — | — | |
мясные,рыбные | 21,96 | 5,3 | — | ||
растительные | 21,54(29,64%) | 12,86(18,06%) | — | — | |


Таблица 28. Средний суточный набор продуктов для детей от 14 до 17 лет
Наименованиепродуктов | Кол-во | Пищевыевещества, г | Калорийность, ккал | ||
белки | жиры | углеводы | |||
Молоко | 650 | 18,2 | 22,75 | 23,2 | 406 |
Творог | 42 | 5,04 | 3,57 | 1,39 | 59,22 |
Сметана | 15 | 0,31 | 4,23 | 0,46 | 42,6 |
Сыр | 25 | 4,67 | 4,08 | 0,45 | 60,75 |
Яйцо | 50 | 4,5 | 4,85 | 0,15 | 63,5 |
Мясои мясопродукты | 190 | 25,08 | 4,94 | — | 148,2 |
Рыба и рыбопродукты | 65 | 7,54 | 0,13 | — | 27,72 |
Хлебпшеничный | 250 | 19,05 | 3 | 122,5 | 608,25 |
» ржаной | 85 | 4,25 | 0,85 | 36,12 | 173,4 |
Мука пшеничная | 15 | 1,4 | 0,15 | 11,45 | 52,5 |
» картофельная | 8 | 0,06 | — | 6,18 | 26,47 |
Крупа гречневая | 8 | 0,94 | 0,14 | 5,04 | 25,6 |
» манная | 4 | 0,29 | 0,02 | 2,8 | 13,2 |
» рисовая | 12 | 0,8 | 0,11 | 8,53 | 39 |
» геркулес | 4 | 0,43 | 0,25 | 2,5 | 1-1 |
» пшено | 12 | 1,2 | 0,26 | 7,85 | 39,6 |
Макароны | 20 | 1,86 | 0,17 | 14,18 | (17,2 |
Сладости | 20 | 1 | 2 | 13,4 | 77,6 |
Сахар | 60 | — | — | 57,3 | 234 |
Картофель | 350 | 4,2 | — | 49 | 218,12 |
Овощиразные | 350 | 3,64 | — | 15,4 | 78 |
Фруктысвежие | 200 | 0,4 | — | 20,2 | 84,46 |
» сухие | 10 | 0,2 | — | 10,1 | 42,23 |
Маслосливочное | 45 | 0,16 | 35,3 | 0,22 | 330 |
» растительное | 17 | — | 16,95 | — | 148,3:) |
Поваренная соль | 10 | — | — | — | — |
Чай | 0,2 | — | — | — | — |
Кофе | 3 | — | — | — | — |
Какао | 1 | 0,2 | 0,19 | 0,38 | 4,1 |
Всего: | 105,42 | 102,98 | 408,8 | 3084 | |
Изних продукты: | |||||
молочные | 28,38 | 69,93 | — | — | |
мясные,рыбные | 37,12 | 9,92 | — | — | |
растительные | 39,92(37,87%) | 23,13(22,46%) | |||
Глава 14
РАССТРОЙСТВА ПИЩЕВАРЕНИЯ
Соблюдение режима питания, недопущение однообразного питания, пищевых излишеств, еды в сухомятку, второпях, отвлечение во время еды может предупредить развитие желудочно-кишечных расстройств у детей. Отмечалось, что при составлении рационов должны учитываться возрастные особенности детей, общее их состояние, индивидуальные особенности роста и развития. Для каждого возраста должен использоваться определенный набор пищевых продуктов. Выполнение этих правил особенно важно для ослабленных детей с признаками функциональных нарушений со стороны желудочно-кишечного тракта, с семейным предрасположением к язвенной болезни желудка, двенадцатиперстной кишки и другим заболеваниям органов пищеварения.
Педиатрам нередко приходится встречаться с острыми заболеваниями желудка и кишечника у детей, причина которых— прием недоброкачественной пищи, пищевое отравление. Что же касается хронических заболеваний органов пищеварения, то до недавнего времени в педиатрии считалось, что такие хронические заболевания, как язва желудка, язва двенадцатиперстной кишки, панкреатит, холецистит, не свойственны детскому возрасту. Однако в последние годы стало известно, что эти болезни у детей не столь уже редкие.
Каковы же причины этих заболеваний? Причины, вызывающие их, многоообразны, и нет возможности все перечислить. Поэтому ограничимся рассмотрением некоторых и, в частности, таких, как недостаточное пережевывание пищи, быстрая еда, частое употребление слишком холодной или горячей пищи, однообразное питание с недостаточным содержанием в пище витаминов. За исключением недоброкачественной пищи, все перечисленные факторы при длительном действии способны оказывать вредное влияние на органы пищеварения. В частности, слишком продолжительные промежутки времени между приемом пищи, которые нередко встречаются у школьников, могут повлечь последствия. Дело в том, что во время длительного перерыва в приеме пищи в пустом желудке появляется 2—3-часовая желудочная секреция, выделяемый при этом желудочный сок обладает высокой кислотностью и оказывает раздражающее действие на слизистую оболочку желудка, в результате чего могут возникнуть такие заболевания, как хронический гастрит и язвенная болезнь.
К числу причин может быть отнесен ряд факторов, способствующих развитию невротических состояний, следствием которых могут быть нарушения деятельности пищеварительных органов (И. Т. Курцин, С. С. Полты-рсв и др.). Что же касается причин возникновения неврозов, то к ним обычно относят психические травмы, отрицательные эмоции, эндокринные расстройства, интоксикации, вызывающие перенапряжение нервных процессов, истощение центральной нервной системы.
Поражения органов пищеварения у детей могут быть обусловлены перенесенными заболеваниями инфекционной природы (грипп, скарлатина, корь, болезнь Боткина, хронический тонзиллит и др.), а также и острыми желудочно-кишечными заболеваниями инфекционной природы (острые гастриты, гастро-дуодениты, гастро-энтериты и др.). Не до конца проведенное лечение может способствовать переходу воспалительного процесса из острого в подострое, а затем и в хроническое течение.
Общие признаки нарушений деятельности пищеварительной системы. Некоторые признаки нарушений деятельности пищеварительного аппарата встречаются при большинстве желудочно-кишечных заболеваний и при этом в самых различных сочетаниях. Таковы: изжога, тошнота, рвота, запор, понос, понижение или потеря аппетита, извращение вкуса и аппетита, боли, отрыжка и др.
Изжога — чувство жжения в нижнем отделе пищевода. Наблюдается при повышенном содержании в желудочном соке соляной кислоты, но может возникать и при нормальной или пониженной кислотности. В этих случаях обусловливается повышенным содержанием в желудочном содержимом органических кислот, например молочной.
Отрыжка — непроизвольное выбрасывание воздуха из желудка в рот. Иногда происходит выбрасывание из желудка пищевых частиц, желудочного сока, желчи. Отрыжка воздухом обычно является следствием усиленного газообразования в желудке (усиленное брожение), спазма пилорического сфинктера, в связи с чем происходит длительная задержка пищи в желудке. Механизм отрыжки рефлекторный. Начальное звено этого рефлекса — рецепторы желудка, подвергающиеся действию повышенного давления или силыюраздражающих веществ. Тошнота — чувство давления в подложечной области с одновременным неприятным ощущением во рту. Тошнота почти всегда возникает перед рвотой, но не всегда она сопровождается ею. Иногда наступает при виде неприятной пищи и восприятии неприятных запахов.
Рвота — непроизвольное выбрасывание желудочного содержимого через рот наружу. Это сложный рефлекторный акт, центр которого расположен в продолговатом мозгу. Во время рвоты происходят закрытие привратника, сокращение пилорического отдела желудка, раскрытие кардиального сфинктера и появление антиперистальтики. Осуществлению рвоты способствует сокращение брюшного пресса, обусловливающее повышение внутри-брюшного давления. В момент рвоты опускается надгортанник с одновременным поднятием гортани и закрытием голосовой щели, что предотвращает попадание рвотных масс в дыхательные пути. Проникновению этих масс в полость носа препятствует поднятие мягкого нёба. Рвота иногда сопровождается антиперистальтикой кишок, что при открытом пилорическом сфинктере может обусловить примешивание к рвотным массам кишечного содержимого (в том числе и желчи). Следует отметить, что причиной рвоты могут быть и нежелудочные факторы (поражение брюшины, заболевания мозговых оболочек и мозга и др.).
Понос — расстройство функции кишечника, выражающееся в появлении жидких и обычно частых испражнений. При поносе наблюдается усиленная перистальтика кишок, нарушения секреторной, ферментовыделительной и всасывательной функций кишечника. Последствием стойких поносов является обезвоживание организма, общее голодание вплоть до истощения. Причинами поносов могут быть регуляторные расстройства пищеварительной системы, наличие воспалительного процесса, токсины а также продукты гниения. При поносах, рвоте может иметь место в связи с обезвоживанием организма ощущение жажды. В механизме ее возникновения важная роль принадлежит центральной нервной системе и, видимо, так называемому центру жажды, расположенному в коре головного мозга (И. Н. Журавлев).
Запор — длительная задержка стула. Причины, вызывающие его, различные: слишком скудная и бедная остатками пища, нарушение регуляции тонуса мускулатуры кишок (атония или спастические сокращения). Стойкие запоры отрицательно влияют на организм, так как при этом усиленно протекают в кишечнике (особенно толстом) гнилостные процессы с образованием индола, скатола, крезола и других ядовитых веществ. Длительное поступление из кишечника в печень избыточного количества этих ядов может нарушать ее барьерную функцию и тогда неизбежна интоксикация.
Метеоризм — вздутие, пучение желудка и главным образом кишечника газами. Возникает вследствие усиления в кишках процессов брожения и гниения. Развитию метеоризма способствует ослабление перистальтики кишок. При метеоризме наблюдается резко выраженное торможение секреторной деятельности главных пищеварительных желез (за исключением слюнных, функция которых усиливается), учащается пульс, к тому же он становится слабым; в начале артериальное давление повышается, а затем падает. Как правило, он сопровождается болями в области живота вследствие сдавления сосудов, питающих стенку кишок, а также чрезмерного раздражения рецепторов, заложенных в стенках кишок. Следует иметь в виду, что боли наблюдаются не только при метеоризме, но и при многих желудочно-кишечных заболеваниях и заболеваниях желчных путей. Часто источником их могут быть спастические сокращения гладкой мускулатуры желудка, кишок, желчного пузыря и т. д. В настоящее время нет достаточных оснований для признания наличия специальных «болевых» рецепторов в стенках пищеварительного тракта. Возможно, что любые рецепторы, подвергаясь сильным воздействиям, могут посылать в центральную нервную систему импульсы, под влиянием которых могут возникать болевые ощущения (В. Н. Черниговский). Однако независимо от причины возникающие стойкие и сильные болевые ощущения оказывают отрицательное влияние на состояние центральной нервной системы и ее высшего отдела — коры головного мозга.
Понижение и извращение вкуса и аппетита. Эти изменения часто встречаются у больных с заболеваниями желудочно-кишечного тракта. Извращение вкуса проявляется в виде неприятного ощущения при приеме пищи, что сопряжено с изменением функционального состояния вкусового анализатора под влиянием импульсов, поступающих из пораженного органа. Изменения аппетита могут быть как в сторону повышения, так и понижения, иногда наблюдается полная его потеря. Все изменения обычно являются следствием нарушения функционального состояния пищевого центра.
Функциональное состояние пищеварительной системы
при патологии пищеварительных органов
Особого внимания заслуживают болезни зубов, часто встречающиеся у детей. Одним из частых признаков поражений зубов является так называемая зубная боль, обычно причиняющая немалые страдания ребенку. Нельзя не учитывать и того обстоятельства, что наличие в кариозных зубах хронической инфекции может быть причиной развития аллергических состояний, расстройств деятельности внутренних органов и систем организма.
М. П. Жаков считает, что в ряде случаев лечение или удаление пораженных зубов улучшает течение некоторых заболеваний сердечно-сосудистой системы, почек и других органов. Экспериментальные данные свидетельствуют о том, что раздражение пульпы зуба может вызвать у подопытных животных ряд общих нарушений в организме: воспаление десен, облысение, одышку, двигательные расстройства и пр. Исследователи наблюдали у собак расстройства деятельности главных пищеварительных желез и мускулатуры желудочно-кишечного тракта, а в некоторых случаях и различного характера структурные изменения. Учитывая это, нетрудно убедиться в том, что своевременное обращение к стоматологу является обязательным во избежание возможных последствий.
Одним из распространенных заболеваний среди детей является воспаление слизистой оболочки рта — стоматит (острый или хронический, язвенный или катаральный). Стоматит сопровождается болевыми ощущениями при приеме пищи, а также изменениями в общем состоянии ребенка. Что же касается состояния пищеварения при стоматитах, то оно еще недостаточно изучено.
В эксперименте на собаках стоматиты воспроизводились обычно путем воздействия сильнораздражающими веществами на слизистую оболочку ротовой полости либо однократно, либо многократно на протяжении одного или более месяцев. Исследования показали, что катаральные стоматиты сопровождаются нарушениями секреторной деятельности слюнных желез, причем чаще наблюдается усиление слюноотделения при одновременном изменении состава отделяемой слюны (снижение сухого остатка). Такой эффект может иметь защитно-приспособительное значение, если учесть наличие в слюне фермента лизоцима, обладающего высоким бактерицидным свойством. С переходом катарального стоматита в язвенный у животных наступает стойкое и резкое понижение слюноотделения и повышение содержания в слюне сухого остатка. Заболевание сопровождается и другими нарушениями в деятельности органов пищеварения. В частности, изменяется двигательная способность пищевода (появляются спастические сокращения), возникает нарушение желудочной секреции, причем чаще всего оно характеризуется угнетением сокоотделения (снижение уровня желудочной секреции и понижение кислотности желудочного сока). При стоматитах у животных наблюдается нарушение желчеотделения, причем при катаральном стоматите образование желчи усиливается, а при язвенном — угнетается. С наступлением заживления слизистой рта желчеотделение восстанавливается. Проводимое лечение стимулирует внешнесекре-торную деятельность печени (С. А. Хачатрян).
Нарушения деятельности слюнных желез наблюдаются при различных заболеваниях органов пищеварения, причем могут характеризоваться усилением и ослаблением слюноотделения. В последнем случае больные ощущают сухость во рту, испытывают затруднения при глотании. Часто изменения слюноотделения сочетаются с изменением отделения слизистыми железами слизи. Механизм развития этих расстройств рефлекторный. Усиление или понижение деятельности слюнных желез можно объяснить изменением функционального состояния слюноотделительного центра, расположенного в продолговатом мозгу и имеющего представительства в других отделах центральной нервной системы. Изменение состояния слюноотделительного центра может быть следствием поступающих к нему «патологических» импульсов из очага поражения.
Нарушения сокоотделения в желудке, выявляемые при специальном зондировании, не всегда указывают на наличие поражений пищеварительного тракта. Они могут встречаться при заболеваниях нервной и эндокринной систем, мочевыделительных органов и т. д. Нарушения могут характеризоваться снижением желудочной секреции и качественными изменениями желудочного сока (повышение или понижение кислотности, понижение активности ферментов и в особенности пепсина). Одним из проявлений секреторных расстройств является извращение динамики сокоотделения. К наиболее тяжелым нарушениям работы желудка относится ахилия — отсутствие отделения кислого желудочного сока, причем в одних случаях она связана с угнетением деятельности желез желудка, в другом — с атрофией железистого аппарата желудка (необратимые органические, структурные изменения слизистой). К секреторным расстройствам желудка относят и нарушения деятельности слизистых желез, расположенных в его слизистой оболочке. Часто при поражении желудка количество отделяемой слизи повышается, что снижает кислотность желудочного сока.
Расстройства моторики и эвакуации взаимосвязаны. С нарушением моторики желудка изменяется и скорость опорожнения его. Нарушения моторики обычно обусловлены изменением тонуса гладкой мускулатуры, которое может проявляться в виде повышения, понижения или полной потери тонуса (атония). К двигательным расстройствам относятся спазмы гладкой мускулатуры. Эвакуаторные нарушения могут проявляться в форме ускоренного или замедленного опорожнения желудка. В частности, при спазме пилорического сфинктера, сужении его просвета рубцовой тканью или опухолью содержимое может длительно задерживаться в желудке. Немаловажную роль может играть и повышенная кислотность желудочного сока. Ускоренная эвакуация химуса из желудка отрицательно влияет на процессы, происходящие в кишечнике.
Расстройства кишечного пищеварения встречаются часто и могут быть обусловлены нарушениями желчеотделения и желчевыделения, отделения поджелудочного и кишечного соков, нарушениями двигательной и секреторной функций желудка. Нарушения желчеотделения могут возникать, например, под влиянием закупорки протоков камнями, сужения желчных ходов, сдавления их опухолями, двигательных расстройств желчных путей. Недостаточное или полное прекращение поступления желчи в тонкий кишечник тяжело отражается на ходе кишечного пищеварения и всасывания (особенно жира). Постепенно происходит ослабление перистальтики в толстом кишечнике, что способствует развитию запоров. Следует упомянуть, что в отсутствии желчи понижается всасывание в кишечнике витаминов (в частности, витамина К), играющих важную роль в обмене веществ и ряде функций организма.
Функциональная недостаточность поджелудочной железы встречается у детей довольно часто, в том числе у детей раннего возраста. Она проявляется как в уменьшении, так и в полном прекращении поступления поджелудочного сока в кишечник, а также снижением его ферментативной активности. Это может быть вследствие поражения поджелудочной железы (панкреатит), закупорки протока железы или сдавления его опухолью. При недостаточности поджелудочной железы кишечное пищеварение страдает очень резко, так как в поджелудочном соке содержатся все ферменты, способные расщеплять углеводы, белки и жиры.
Возможны расстройства деятельности кишечных желез, которые могут проявляться чаще всего в уменьшении количества отделяемого сока и уменьшении содержания в нем ферментов. Все это, естественно, в какой-то мере отражается па интенсивности кишечного пищеварения. В частности, у детей вследствие недостатка в кишечном содержимом ферментов инвертазы или лактазы страдает усвоение молочного сахара. В меньшей степени страдает расщепление белков и жиров, так как компенсация возможна за счет усиления выделения соответствующих ферментов поджелудочной железой.
Нарушения всасывания обычно бывают от недостаточно полного расщепления составных частей пищи. При воспалительных процессах в тонкой кишке всасывание может изменяться как в сторону усиления, так и ослабления. Длительное и стойкое ослабление всасывания в тонкой кишке неизбежно приводит к истощению организма, снижению массы тела. В случае же повышенной проницаемости слизистой оболочки тонкой кишки, что часто имеет место при воспалительных процессах в кишечнике, в кровь могут проникать продукты неполного расщепления пищи, ядовитые вещества (токсины микробного и гельминтозного происхождения), вызывающие кишечную интоксикацию.
Необходимо иметь в виду, что при расстройствах пищеварения обычно страдает не одна какая-либо функция пищеварительного органа, а вся его деятельность в целом. Больше того, расстройство деятельности одного из пищеварительных органов нередко сопровождается нарушением в деятельности других органов. Это и попятно, если учесть, что пищеварительная система — единая и все ее части взаимосвязаны, а функции взаимообусловлены.
Экспериментальными исследованиями выявлен механизм развития пищеварительных расстройств. Первые и наиболее убедительные данные относительно механизма развития секреторных расстройств при патологии желудка были получены в конце XIX — начале XX в. И. П. Павловым и его школой. В частности, было показано, что при гастрите, воспроизведенном в большом желудке, нарушения секреции обнаруживались и в павловском изолированном желудочке. Больше того, секреторные расстройства наблюдались и в изолированном желудочке по Гейденгайпу (с выключенной парасимпатической иннервацией). Это указывает на участие в механизме развития секреторных расстройств не только нервной системы, но и гуморальных факторов.
Нервно-гуморальный механизм лежит и в основе развития секреторных нарушений в кишечнике. При повреждении, локализующемся в одном участке кишечника (прямая и слепая кишки и т. д.), нарушения секреции регистрируются в других участках его (С. С. Пол-тырев с сотрудниками). Важную роль в осуществлении патологических влияний с одних участков пищеварительного тракта на другие играет кора головного мозга (Н. А. Рощина, Г. В. Николаева). Установлено, что и у собак с удаленной корой головного мозга указанные патологические влияния возможны, но проявляются они иначе, чем до декортикации. В этом мы видим выключение коррегирующего влияния коры мозга. Коррекция резко страдает и у животных с экспериментальным неврозом.
Не все происходящие изменения в деятельности пищеварительного аппарата можно отнести к патологическим сдвигам. Некоторые из них могут быть защитными приспособлениями, в частности рвота, усиление отделения слизи, выделения с пищеварительными соками продуктов азотистого обмена (шлаков), бактерицидное действие лизоцима слюны и соляной кислоты желудочного сока, обезвреживание в печени ядовитых веществ, образующихся в толстом кишечнике при гниении, избирательная способность слизистой оболочки кишок к всасыванию некоторых составных частей кишечного содержимого, изменение аппетита. Все эти защитные приспособления являются результатом эволюции. У высших животных они более совершенны, чем у животных, стоящих на более низких ступенях развития.
Для приведения. в действие ряда защитно-приспособительных и компенсаторных реакций, защитных приборов важное значение имеет раздражение рецепторов пищеварительного тракта, а возникающие при этом рефлекторные акты осуществляются при непосредственном участии центральной нервной системы. Имеется немало фактов, указывающих на кортикальный механизм мобилизации мер защиты (И. Т. Курцип, С. С. Полтырев и др.). Известно, что у собак с экспериментальными неврозами проявление-защитных реакций на действие вредных агентов утрачивает свою закономерность и поэтому заболевание протекает тяжелее и иногда заканчивается гибелью животного. Кортикальный механизм находит подтверждение и в том, что проявление защитной реакции может быть вызвано на условный (индифферентный) раздражитель, если он (звонок, свисток, вспышка света и др.) несколько раз приводился в сочетание с непосредственным действием болезнетворного агента (например, условнорефлекторная рвота, слюноотделение и др.).
Необходимо иметь в виду, что при некоторых условиях защитные реакции могут оказаться недостаточными для предотвращения развития заболевания. Иногда же они на каком-то этапе заболевания могут выключаться и тем самым осложнить его течение.
Некоторые заболевания желудочно-кишечного тракта у детей
Рассмотрим некоторые заболевания желудочно-кишечного тракта у детей.
Гастрит — воспалительное заболевание слизистой оболочки желудка, которое может возникать под влиянием различных причин. Среди них существенную роль играют длительные нарушения качества и режима питания: продукты, механически и химически раздражающие слизистую оболочку (копчености, пряности, маринады и др.), длительно и трудно перевариваемые продукты (жирная свинина и баранина, жирные колбасы, гуси, утки), недостаток в пище витаминов (А, тиамина, рибофлавина, аскорбиновой и никотиновой кислот). Температура принимаемой пищи и питья также имеет значение: горячая вода 60—80° С повреждает слизистую оболочку желудка. Имеет значение еда в сухомятку, на ходу, несоблюдание режима при приеме пищи. Перечисленные факторы относятся к внешним — экзогенным. Однако немаловажную роль играют и внутренние факторы — эндогенные, среди которых одно из первых мест принадлежит нарушениям деятельности нервной системы (центральной, вегетативной), а также рефлекторным воздействиям с различных пораженных органов (кишечника, желчного пузыря и т. д.). Они могут вызывать изменения секреторной деятельности желудочных желез, а в дальнейшем служить причиной развития структурных изменений в слизистой оболочке желудка. Возможен переход острого гастрита в подострую и хроническую формы с постепенно прогрессирующими морфологическими изменениями в стенке желудка (С. М. Рысс).
По характеру структурных изменений хронический гастрит делят на поверхностный с поражением желудочных желез, но без атрофии их, атрофический и гипертрофический; по функциональному признаку — на гастрит с нормальной секреторной функцией, усиленной функцией, с умеренно и резко выраженной секреторной недостаточностью, о чем судят по состоянию кислотности желудочного сока и его переваривающей способности.
Врачебная практика показывает, что хронический гастрит может протекать бессимптомно. Следует, однако, учесть, что в силу различных причин заболевание может обостриться. У детей важным симптомом является диспепсия, которая проявляется дискомфортом: отрыжка, срыгивания, ощущения тяжести в животе, неприятные ощущения во рту, тошнота, снижение, отсутствие или извращение аппетита. Иногда проявляется кишечная диспепсия: ощущение переполнения пищей, метеоризм, усиление перистальтики (урчание в животе), запоры, поносы. Нередко ощущения переполнения и тяжести после еды больными принимаются за боли. Боли при хронических гастритах возникают чаще всего после приема пищи и удерживаются час, а иногда и дольше.
Появление их объясняют растяжением стенок желудка пищей, повышенной возбудимостью рецепторного аппарата желудка.
По наблюдениям М. Б. Коссюры, хронический гастрит развивается у детей дошкольного и школьного возрастов. Среди больных гастритом с пониженной кислотностью преобладали дети, заболевшие в раннем и дошкольном возрасте, а с сохранившейся кислотностью желудочного сока — дети, заболевшие в школьном возрасте. Автор высказывает предположение, что пониженная кислотность у детей раннего возраста может быть иногда врожденной. У детей с хроническим гастритом преобладала повышенная кислотность, вдвое реже пониженная и редко нормальная.
М. Б. Коссюра на основании многолетних клинических наблюдений считает, что следует различать две формы гастрита детей: малосимптомную, с преимущественно легким течением, и более выраженную клиническими проявлениями. При второй форме боли интенсивные, нередко голодные, ночные. У детей отмечается повышенная общая возбудимость. Кислотность желудочного содержимого обычно повышена. При малосимптомной форме заболевание протекает монотонно со слабо выраженными жалобами, возникающими после еды, боли неинтенсивны и непродолжительны.
Длительные наблюдения за больными хроническим гастритом показали, что болезнь нередко принимает затяжное течение, но в ряде случаев возможно полное выздоровление. Огромное значение придается созданию ребенку спокойной обстановки, поддержанию строгого режима дня, предоставлению достаточного времени для сна и пребывания на воздухе. Детей школьного возраста не следует перегружать домашними заданиями. Из лечебных мероприятий первое место отводится соблюдению предписанного врачом режима и диеты, систематическому приему минеральных вод, витаминов, в частности аскорбиновой кислоты.
Дети с хроническим гастритом должны находиться под систематическим наблюдением врача.
Язвенная болезнь — заболевание, характеризующееся образованием язвы в желудке или двенадцатиперстной кишке. С началом широкого применения рентгенологического метода исследования в педиатрической практике у детей стали чаще регистрироваться случаи язвенной болезни. Среди 764 обследованных детей с различными заболеваниями пищеварительной системы М. Б. Коссюра выявила язвенную болезнь у 32 (4%). В возникновении язвенной болезни придается немаловажное значение нервно-психическому фактору: нервным переживаниям, нарушению режима и рациона питания, наследственно-конституциональному фактору. В отдельных случаях болезнь протекает скрыто, без симптомов (у 2/3 обследованных детей). В настоящее время считают, что язвенная болезнь может развиться у детей всех возрастов и разного пола с четырехлетнего до 15-летнего возраста. Высказывается предположение, что язвенная болезнь взрослых в ряде случаев начинается в детском возрасте.
У детей с язвенной болезнью наблюдается пониженный аппетит, некоторая задержка роста. Важнейшим симптомом является боль в животе после еды, но чаще всего голодная или ночная. Нередко боль сочетается с диспепсическими явлениями. Наиболее часто у детей наблюдаются рвота, тошнота, отрыжка, изжога. При язвенной болезни характерна повышенная кислотность желудочного сока, причем нередко стойко удерживающаяся. Только в период выздоровления она начинает снижаться.
Диагноз язвенной болезни обычно основывается на данных клинических и рентгенологических исследований. У больных могут обнаруживаться нарушения функций поджелудочной железы, печени и желчных путей, кишечника, что указывает на функциональную связь разных органов пищеварительной системы.
Больные дети отличаются повышенной нервной возбудимостью, раздражительностью, изменениями со стороны вегетативного отдела нервной системы, наблюдается потливость, замедленный пульс, пониженное артериальное давление, меняется состав крови.
У детей бывают случаи осложнений: сужение привратника, желудочно-кишечное кровотечение, прободение. В связи с этим прибегают к оперативному вмешательству.
Лечение комплексное с применением диеты, медикаментов, физиотерапевтических курортных факторов.
Хронические заболевания поджелудочной железы. В педиатрии считалось, что хронические заболевания поджелудочной железы являются редкостью. Вместе с тем для их возникновения у детей существуют не меньшие предпосылки, чем у взрослых. Это касается анато-мо-физиологических особенностей железы, напряженного обмена веществ у детей, определяющего повышенные требования к разнообразной ее деятельности. Как указывалось, поражение поджелудочной железы может быть обусловлено различными заболеваниями, перенесенными в детстве.
Л. В. Клименская показала, что наиболее частыми причинами функциональных нарушений вненшесекре-торной функции поджелудочной железы и панкреатита являются язвенная болезнь, воспалительные заболевания желчных путей, хронический гастрит, грубые и длительные нарушения режима и характера питания. Она описала две формы хронического панкреатита: 1) хроническую, повторяющуюся с выраженной болью, 2) латентную. Для первой формы характерны упорные боли в сочетании с диспепсическими проявлениями. Для второй— сначала диспепсические расстройства с нарушением аппетита и извращением вкуса, отрицательным отношением к жирной пище, сладостям, часты рвота и тошнота, в дальнейшем — общее недомогание, разбитость, подавленность, головная боль, прогрессирующее исхудание, неустойчивый стул. Распознать заболевание трудно в связи с тем, что ряд признаков встречается и при других заболеваниях пищеварительной системы.
Поскольку хронический панкреатит часто сочетается с заболеваниями других пищеварительных органов, с нарушениями их деятельности, то проводимое лечение предусматривает воздействие не только на поджелудочную железу, но и на те органы, которые вовлечены в круг патологического процесса. Необходимо отметить, что у детей при панкреатитах может изменяться не только внешнесекреторная функция, но и внутрисекреторная (образование гормонов).
Кишечная интоксикация. У здорового человека в толстом кишечнике всегда имеется разнообразная микрофлора. И. И. Мечников отметил, что между гнилостными микробами и бактериями, вызывающими брожение, существует антагонизм. Доказано, что кишечная палочка тормозит развитие ряда патогенных микробов.
В условиях патологии пищеварительного тракта микрофлора кишечника может оказывать вредное действие на организм, вызывая интоксикацию. Необходимым условием ее возникновения является прежде всего нарушение двигательной функции кишечника — ослабление перистальтики, влекущее за собой запоры, при которых страдает не только моторика, но и секреция пищеварительных соков. Обычно сокоотделение в кишечнике снижается и понижается активность ряда ферментов, вследствие чего нарушаются кишечное пищеварение и всасывание. Все это, естественно, создает весьма благоприятные условия для усиления гнилостных и бродильных процессов. Гнилостные процессы, получив широкое развитие, неизбежно ведут к образованию ядовитых продуктов: скатол, индол, крезол, фенол. Обезвреживание происходит в печени путем окисления и связывания их с глюкуроновой или серной кислотами, в результате чего образуются безвредные соединения, выделяющиеся через почки. Гнилостному брожению подвергаются аминокислоты, образуя амины (гистамин, кадаверин, пут-ресцин и др.).
Поступление в печень в течение продолжительного времени значительных количеств ядовитых веществ, образующихся в кишечнике при гниении белков, может обусловить развитие в паренхиме печени жирового перерождения. Вместе с тем при недостаточности функций печени, снижении ее обезвреживающей способности поступающие из кишечника яды могут стать причиной тяжелого отравления.
У детей причиной кишечной интоксикации нередко являются токсические диспепсии, возникающие под влиянием экзогенной инфекции и путем активации собственных микробов кишечника в условиях перегревания, перенесенных заболеваний, расстройств питания и пр. Большое значение в происхождении, течении и исходе токсической диспепсии принадлежит печени. В ней накапливается много бактериотоксинов, биогенных аминов, органических кислот и чрезвычайно ядовитых соединений ароматического ряда, в связи с чем ее антитоксическая способность подвергается большим испытаниям. Этим создаются условия для «прорыва» печеночного барьера и поступления вредных веществ в общий круг кровообращения. По мере накопления в кишечнике ядовитых веществ они оказывают раздражающее действие на хеморецепторы и рефлекторно вызывают реакции со стороны различных органов и систем организма. И. М. Уткина в опытах на собаках показала, что орошение слизистой оболочки изолированной петли тощей кишки, павловского изолированного желудочка экстрактами из аскарид сопровождается клиническими признаками интоксикации и нарушением секреторной и экскреторной функций желудка. Выключение рецепторов изолированной петли кишки 2%-ным раствором новокаина делают невозможным осуществление таких влияний с кишечника. Орошение экстрактами из аскарид денсрви-рованного изолированного желудочка или денервирован-ной петли кишки не сопровождает развитие картины интоксикации и секреторных расстройств желудка. Эти данные указывают на участие в развитии интоксикации рефлексов с хеморецепторов кишечника.
В различные стадии развития кишечной интоксикации удается наблюдать неодинаковые сдвиги со стороны артериального давления. Так, в начальном периоде нередко наблюдается его повышение, однако в дальнейшем с нарастанием токсикоза оно падает и достигает низкого уровня, причем более низкого, чем до заболевания. Электрокардиографические исследования показывают наличие у животных с экспериментальной кишечной интоксикацией изменений зубцов Р и Т, выражающихся в уплощении, двухфаз'ности и отрицательном направлении этих зубцов, а также в увеличении зубца S и смещении интервала S—Т. Эти отклонения указывают на развитие в миокарде диффузных изменений, на перенапряжение правого желудочка.
Кишечная интоксикация обычно сопровождается снижением почечной функции и особенно фильтрационной способности почек. Очевидно, нарушение клубочко-вой фильтрации — следствие спазма капиллярных петель. Нарушение деятельности почек может отягощать течение интоксикации, так как при этом возможна задержка в организме азотистых шлаков и других ядовитых веществ. В крайне тяжелых случаях интоксикации возможно развитие структурных изменений в почках и при этом даже необратимых.
Нередко аутоинтоксикация сопровождается ацидозом (понижением рН крови). Одним из факторов, обусловливающим его развитие, является снижение активности фермента карбоангидразы, что ведет к накоплению в крови и тканях углекислоты и нарушению тканевого дыхания.
Некоторые изменения, возникающие в деятельности различных органов, носят не патологический, а защитно-компенсаторный характер, разграничить их бывает очень трудно. Например, в разные стадии развития интоксикации обнаруживаются фазовые изменения в функциональном состоянии коры головного мозга: в начальном периоде кратковременная фаза возбуждения (повышенная возбудимость коры мозга устанавливается применением метода электроэнцефалографии), после чего возбуждение сменяется торможением, которое можно до известной степени рассматривать как охранительный процесс.
Причиной ряда расстройств в организме может быть метеоризм как следствие пареза кишечной мускулатуры, ослабления перистальтики.
Клинические наблюдения дают основание утверждать, что аутоинтоксикация у детей протекает более тяжело, чем у взрослых. Это, видимо, объясняется тем, что приспособительные возможности к условиям среды у ребенка раннего возраста значительно меньшие, чем у взрослого; нервно-рефлекторный механизм еще недостаточно совершенен, так как имеется незаконченность морфологического и функционального развития коры головного мозга и неустойчивость функционального состояния подкорковых образований.
Пищеварительные расстройства при патологии некоторых органов и систем
Пищеварительные расстройства не всегда возникают при патологии самой пищеварительной системы.
Пищеварение при нарушениях гормонального баланса в организме
Эндокринные заболевания у детей не являются редкостью. Одни дети страдают заболеваниями щитовидной железы, другие — поджелудочной, третьи — гипофиза и т. д. Довольно часто наблюдается нарушение в работе органов пищеварения при заболеваниях щитовидной железы. Иногда в клинической картине тиреотоксикоза, связанного с повышением гормональной активности щитовидной железы, центральное место занимают желудочно-кишечные расстройства (нарушения аппетита, приступы болей в животе, учащенный кашицеобразный стул, иногда неукротимая рвота и др.). При базедовой болезни резко ускоренно происходит движение по кишечнику содержимого, в связи с чем многие питательные вещества не всасываются. При мик-седеме (функциональная недостаточность щитовидной железы) отмечается сухой и утолщенный язык, исчезновение аппетита, тошнота, запор, метеоризм в связи с ослаблением перистальтики кишечника. Пищеварительные расстройства наблюдаются и при других заболеваниях эндокринной системы, в частности при поражениях надпочечников, гипофиза. Однако эти расстройства функций пищеварительных органов неспецифические, их обычно рассматривают как сопутствующие и возникающие вследствие нарушения в организме нейро-гумо-ральной регуляции.
Пищеварение при почечной недостаточности
Довольно полно изучена секреторная деятельность желудка при почечной недостаточности. Известно, что при остром нефрите желудочная секреция и кислотность желудочного сока снижаются. При хронических заболеваниях почек у больных отмечается стойкое угнетение секреторной деятельности желудка, отсутствие в желудочном соке свободной соляной кислоты. Угнетение желудочных желез сочетается со снижением двигательной активности желудочно-кишечного тракта. Часто наблюдается задержка принятой пищи в желудке. Желудочные железы начинают выделять в полость желудка белок, азотистые шлаки (мочевину, креатинин) и электролиты в повышенном количестве, что может оказывать вредное действие на слизистую оболочку желудка и вызывать не только функциональные, но и структурные нарушения. Все указанные нарушения обнаруживаются у животных с искусственно воспроизведенной патологией почек.
Пищеварительные расстройства возникают и при заболеваниях мочевыводящих путей (мочевого пузыря, мочеточников и мочеиспускательного канала). В частности, при воспалительном процессе в мочевом пузыре (цистите) наблюдается нарушение всех основных функций желудка и тонкой кишки. Они возникают также при острой задержке и в особенности при повторных задержках мочи в мочевом пузыре.
Экспериментальные исследования М.В. Саликовой и других показали, что механизм патологических влияний с мочевыводящих путей — нейро-гуморальный.
Пищеварение при патологии дыхательной системы
У собак с воспроизведенными воспалительными процессами в легких (С. А. Конокотина, А. А. Шарыгин, Т. В. Шахова и др.) выявлены нарушения в работе желудка, кишечника, печени. Были вскрыты механизмы, участвующие в осуществлении патологических влияний с легких и плевры на функции органов пищеварения. Выяснилось, что в них участвуют оба звена: нервный, или рефлекторный, и гуморальный. Имеются доказательства, что к моменту ликвидации очага поражения полного восстановления нарушенных функций не наступает, т. е. восстановление может задержаться. Вместе с тем оно наступит тем быстрее, чем раньше нормализуется состояние регулирующих систем организма и прежде всего центральной нервной системы. Скорость нормализации, естественно, зависит и от защитно-компенсаторных реакций. При комплексном лечении эффект резко возрастает.
ЛИТЕРАТУРА
А.В.Асатиани,А.Н.Бакурадэе Нейро-гуморальные механизмы пищеварительной деятельности. Тбилиси, 1975.
Аршавский И.А. Очерки по возрастной физиологии. М., 1967.
Бабкин Б.П. Секреторный механизм пищеварительных желез. Л., 1960.
Бабский Е.Б. и др. Физиология человека. М., 1972.
П.Г.Богач Механизмы нервной регуляции моторной функции тонкого кишечника. Киев, 1961.
И.А.Булыгин Исследование закономерностей и механизмов интероцептивных рефлексов. Минск, 1959.
Булыгин И. А. Электрофизиологический анализ висцеральных афферентных систем. Минск, 1973.
Быков К. М. Кора головного мозга и внутренние органы. М. — Л., 1947.
Быков К. М,, Курцин И.Т.Кортико-висцеральная патология. Л., 1960.
Виру А. А. Функции коры надпочечников при мышечной деятельности. М., 1977.
Гальперин С. И. Физиологические особенности детей. М., 1965.
Горшкова С. М., Курцин И.Т.Механизмы желчевыделения. Л., 1967.
Грачев И. И. Рефлекторная регуляция лактации. М., 1964.
Губарь В. Л. Физиология и экспериментальная патология желудка. М'., 1970.
Двинянинов Л. И. Работы по физиологии и патологии пищеварения, выполненные в лаборатории И. П. ПавловА.М.— Л., 1961.
Двинянинов Л. И. Физиология и патология желудочно-кишечного тракта. Л., 1966.
Дионесов С. М. Боль и ее влияние на организм человека и животных. М„ 1963.
Жеребцов П. И. Экскреторные процессы в желудочно-кишечном тракте. М., 1957.
Кабанов А. Н., Чабовская А. П. Анатомия, физиология и гигиена детей дошкольного возраста. М„ 1975.
Касьяненко И. В. Роль гормонов коры надпочечников в физиологии и патологии желудка. Киев, 1968.
Климов П. К. Механизмы регуляции функций желчевыделительной системы. Л., 1969.
Колосов Н. Г. Иннервация пищеварительного трактА.М.— Л., 1962.
Космическая биология и медицина. М., 1975.
Коссюра М. Б. Болезни желудка у детей. М., 1968.
Курцин И.Т.Гормоны пищеварительной системы. Л., 1962.
Курцин И.Т.Кровоснабжение главных пищеварительных желез. Л., 1976.
Курцин И.Т.Механорецепторы желудка и работа пищеварительного аппарата. М- — Л., 1952.
Лебедев Н. Н. Физиология и патология периодической деятельности пищеварительного тракта. Л., 1967.
Леонтьева Н. П., Мартова К- В., Каплун Э. Г. Анатомия и физиология детского организма. Ч. 1. М., 1976.
Леонтьева Н. Н., Маринова К- В. Анатомия и физиология детского организма. Ч. 2. М., 1976.
Матросова Е. М. Двигательная деятельность желудка и ее связь с секрецией желудочного сока. Л., 1964.
Маркосян А. А. Вопросы возрастной физиологии. М., 1974.
Маянская К- А. Функциональные взаимосвязи органов пищеварения. Л., 1970.
Могендович М. Р. Рефлекторное взаимодействие локомоторной и висцеральной систем. Л., 1957.
Мусящикова С. С, Черниговский В. Н. Кортикальное и субкортикальное представительства висцеральных систем. Л., 1973.
Орбели Л. А. Адаптационно-трофическая функция нервной системы. Т. 2. Л„ 1962.
Основы морфологии и физиологии организма детей и подростков. М., 1969.
Проблемы физиологии гипоталамуса. Гииоталамическая регуляция вегетативных функций. Киев, 1969.
Павлов И. П. Полное собрание трудов. Т. 2. М. — Л., 1946.
Полтырев С.С.Методика проведения опытов по физиологии животных. Ярославль, 1977.
Полтырев С.С.Патологическая физиология пищеварения. Руководство по патологической физиологиИ.Т.4. М., 1966.
Разенков И. П. Влияние пониженного барометрического давления на процессы пищеварения. М., 1948.
Свистун Т. И. Секреция пищеварительных желез во время мышечной деятельности. Киев, 1975.
Селье Г. Очерки об адаптационном синдроме. М., 1960. Соловьев А. В. Новые данные о секреторной функции желудка и поджелудочной железы. Л., 1959.
Сорокин А. П., Стельников Г. В., Вазин А. П. Адаптация и управление свойствами организма. М., 1977.
Скляров Я- П. Секреторная работоспособность главных пищеварительных желез. Киев, 1958.
Уголев А.М.Мембранное пищеварение. Полисубстратные процессы. Организация и регуляция. Л., 1972.
Уголев А.М.Физиология и патология пристеночного пищеварения. Л., 1967.
Уголев А.М.Энтериновая (кишечная гормональная) система. Л., 1978.
Файтельберг Р. О. Всасывание в желудочно-кишечном тракте. М„ 1976.
Физиология пищеварения. Руководство по физиологии. Л., 1974.
Физиология человека/Под ред. Н. В. Зимкина. М., 1975.
Хрипкова А. Г. Взаимодействие симпатического и блуждающего нервов в секреторной деятельности желудка. Ростов-на-Дону, 1962.
Черниговский В, Н. Интероцепторы. М., 1960.
Шлыгин Г. К. Ферменты кишечника в норме и патологии. Л., 1967.
А.Ю.Юнусов, Г.Ф.Коротько Функции органов пищеварения в жарком климате. Ташкент,1962.
Яковлев П. Н. Биохимия спорта. М., 1974.